Витамины компоненты клеточных мембран
Клетка — это не только жидкость, ферменты и другие вещества, но и высокоорганизованные структуры, называемые внутриклеточными органеллами. Органеллы для клетки не менее важны, чем ее химические составляющие. Так, при отсутствии таких органелл, как митохондрии, запас энергии, извлеченной из питательных веществ, сразу же уменьшится на 95%.
Большинство органелл в клетке покрыты мембранами, состоящими в основном из липидов и белков. Различают мембраны клеток, эндоплазматического ретикулума, митохондрий, лизосом, аппарата Гольджи.
Липиды нерастворимы в воде, поэтому в клетке они создают барьер, препятствующий движению воды и водорастворимых веществ из одного компартмента в другой. Молекулы белка, однако, делают мембрану проницаемой для разных веществ с помощью специализированных структур, называемых порами. Множество других мембранных белков являются ферментами, катализирующими многочисленные химические реакции, которые будут рассмотрены в следующих главах.
Клеточная (или плазматическая) мембрана представляет собой тонкую, гибкую и эластичную структуру толщиной всего 7,5-10 нм. Она состоит в основном из белков и липидов. Примерное соотношение ее компонентов таково: белки — 55%, фосфолипиды — 25%, холестерол — 13%, другие липиды — 4%, углеводы — 3%.
Липидный слой клеточной мембраны препятавует проникновению воды. Основу мембраны составляет липидный бислой — тонкая липидная пленка, состоящая из двух монослоев и полностью покрывающая клетку. По всей мембране располагаются белки в виде крупных глобул.
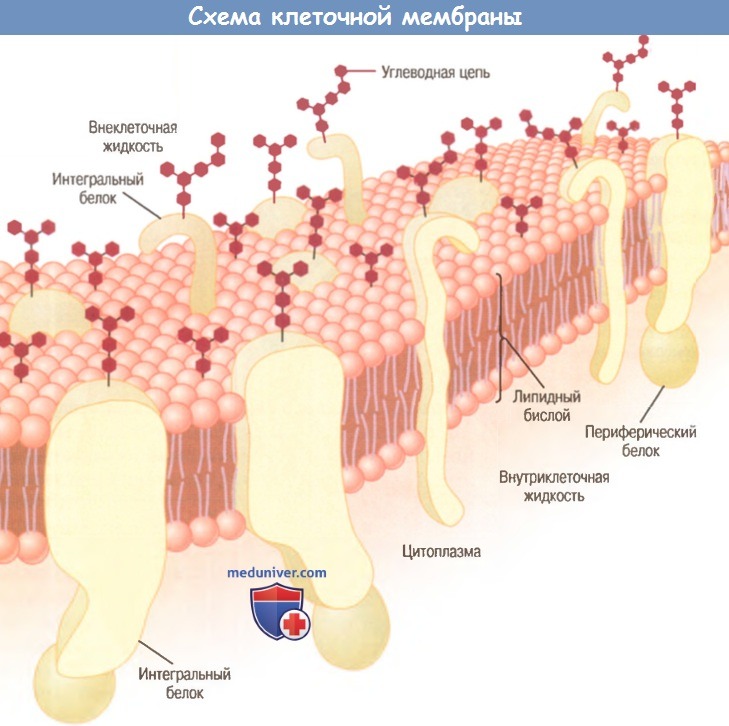 Схематичное изображение клеточной мембраны, отражающее ее основные элементы
Схематичное изображение клеточной мембраны, отражающее ее основные элементы
— фосфолипидный бислой и большое количество молекул белка, выступающих над поверхностью мембраны.
Углеводные цепочки прикреплены к белкам на наружной поверхности
и к дополнительным молекулам белка внутри клетки (на рисунке это не показано).
Липидный бислой состоит главным образом из молекул фосфолипидов. Один конец такой молекулы является гидрофильным, т.е. растворимым в воде (на нем расположена фосфатная группа), другой — гидрофобным, т.е. растворимым только в жирах (на нем находится жирная кислота).
Благодаря тому, что гидрофобная часть молекулы фосфолипида отталкивает воду, но притягивается к подобным частям таких же молекул, фосфолипиды имеют природное свойство прикрепляться друг к другу в толще мембраны, как показано на рис. 2-3. Гидрофильная часть с фосфатной группой образует две мембранные поверхности: наружную, которая контактирует с внеклеточной жидкостью, и внутреннюю, которая контактирует с внутриклеточной жидкостью.
Середина липидного слоя непроницаема для ионов и водных растворов глюкозы и мочевины. Жирорастворимые вещества, включая кислород, углекислый газ, алкоголь, напротив, легко проникают через эту область мембраны.
Молекулы холестерола, входящего в состав мембраны, по природе также относятся к липидам, поскольку их стероидная группировка обладает высокой растворимостью в жирах. Эти молекулы как бы растворены в липидном бислое. Их главное назначение — регуляция проницаемости (или непроницаемости) мембран для водорастворимых компонентов жидких сред организма. Кроме того, холестерол — основной регулятор вязкости мембраны.
Белки клеточных мембран. На рисунке в липидном бислое видны глобулярные частицы — это мембранные белки, большинство которых являются гликопротеинами. Различают два типа мембранных белков: (1) интегральные, которые пронизывают мембрану насквозь; (2) периферические, которые выступают только над одной ее поверхностью, не достигая другой.
Многие интегральные белки формируют каналы (или поры), через которые во внутри- и внеклеточную жидкость могут диффундировать вода и водорастворимые вещества, особенно ионы. Благодаря избирательности действия каналов одни вещества диффундируют лучше других.
Другие интегральные белки функционируют как белки-переносчики, осуществляя транспорт веществ, для которых липидный бислой непроницаем. Иногда белки-переносчики действуют в направлении, противоположном диффузии, такой транспорт называют активным. Некоторые интегральные белки являются ферментами.
Интегральные белки мембраны могут служить также рецепторами для водорастворимых веществ, включая пептидные гормоны, поскольку мембрана для них непроницаема. Взаимодействие белка-рецептора с определенным лигандом приводит к конформационным изменениям молекулы белка, что, в свою очередь, стимулирует ферментативную активность внутриклеточного сегмента белковой молекулы или передачу сигнала от рецептора внутрь клетки с помощью вторичного посредника. Таким образом, интегральные белки, встроенные в клеточную мембрану, вовлекают ее в процесс передачи информации о внешней среде внутрь клетки.
Молекулы периферических мембранных белков часто бывают связаны с интегральными белками. Большинство периферических белков являются ферментами или играют роль диспетчера транспорта веществ через мембранные поры.
Источник
БИОЛОГИ́ЧЕСКИЕ МЕМБРА́НЫ
В книжной версии
Том 3. Москва, 2005, стр. 499-500
Скопировать библиографическую ссылку:





БИОЛОГИ́ЧЕСКИЕ МЕМБРА́НЫ (лат. membrana – кожица, оболочка, перепонка), структуры, ограничивающие содержимое клеток (клеточная, или плазматическая, мембрана, плазмалемма) и внутриклеточных органелл. У прокариот имеется только клеточная мембрана, в большинстве случаев окружённая клеточной стенкой . У эукариот мембраной окружена не только клетка, но и ядро, а также митохондрии, лизосомы, пероксисомы, секреторные гранулы, эндосомы, у растений ещё – хлоропласты и вакуоли; мембраны образуют также разветвлённую сеть эндоплазматического ретикулума и комплекса Гольджи. Митохондрии, хлоропласты и ядра окружены двумя мембранами, а внутри хлоропластов имеется ещё один тип мембран, формирующих тилакоиды. У животных к клеточной мембране снаружи примыкает гликопротеиновый комплекс – гликокаликс , у растений – клеточная стенка. Толщина мембран варьирует от 6 до 10 нм.
Источник
МЕМБРАНЫ БИОЛОГИЧЕСКИЕ
Мембраны биологические (лат. membrana кожица, оболочка) — функционально активные поверхностные структуры клеток толщиной в несколько молекулярных слоев, ограничивающие цитоплазму и большинство внутриклеточных структур, а также образующие единую внутриклеточную систему канальцев, складок и замкнутых полостей.
Мембрану, ограничивающую цитоплазму клетки снаружи, называют плазматической или цитоплазматической мембраной, оболочкой клетки или плазмолеммой. Название внутриклеточных (субклеточных) мембран обычно происходит от названия ограничиваемых или образуемых ими субклеточных структур. Напр., различают митохондриальную, ядерную и лизосомную мембраны, мембраны комплекса Гольджи, эндоплазматического ретикулума, саркоплазматического ретикулума и т. д. (см. Клетка).
Толщина Мембраны биологической ок. 10 нм. однако вследствие сравнительно плотной упаковки основных молекулярных компонентов (белков и липидов) в Мембране биологической, а также большой общей площади клеточных мембран они составляют обычно более половины всей массы клетки (в пересчете на сухой вес). Функции, выполняемые Мембранами биологическими, чрезвычайно важны и разнообразны: формирование клеточных структур, поддержание внутриклеточного гомеостаза, участие в процессе возбуждения и проведения нервного импульса, фото-, механо- и хеморецепция, всасывание, секреция и газообмен, тканевое дыхание, запасание и трансформация энергии и т. д. Всем этим определяется общебиологическое значение М. б. как универсальной и доминирующей формы структурной и функциональной организации живой материи.
Огромное значение Мембран биологических определяется важностью перечисленных выше функций в процессах нормальной жизнедеятельности, а также многообразием заболеваний и патологических состояний, возникающих при нарушениях функций М. б. и проявляющихся на различных уровнях организации— от клетки и ее субклеточных систем до тканей, органов и организма в целом. Подавляющее число известных заболеваний человека и животных являются либо прямым следствием нарушений мембран, либо процессами и состояниями, в большей или меньшей степени сопряженными с ними.
Содержание
История
Термин «мембрана» для обозначения поверхностных образований клетки был введен нем. исследователями Молем (H. Mohl, 1851) при описании плазмолиза клеток растений и Негели (С. W. Nageli, 1855) при изучении механизма осмотических явлений и проникновения в клетки красителей. В 1877 г. Пфеффер (W. F. Ph. Pfeffer) обосновал существование клеточной мембраны, продемонстрировав общность осмотических свойств клеток и осмометров, образованных искусственными полупроницаемыми мембранами. В 80-х гг. 19 в. Х. де Фрис обнаружил, что цитоплазма растительных клеток заключена между двумя мембранами — плазмолеммой и тонопластом.
Первые указания на то, что в состав мембраны клетки входят липиды, получил Овертон (Е. Overton, 1895—1902), к-рый обнаружил прямую связь между растворимостью многих веществ в липидах и их скоростью проникновения в клетку. В 1925 г. Гортер (E. Gorter) и Гренделл (F. Grendell) экспериментально показали, что в мембране эритроцитов количество липидов достаточно для построения двойного непрерывного слоя. Это позволило им высказать предположение, что поверхностная мембрана эритроцита содержит бимолекулярный липидный слой. Примерно в это же время Фрик (H. Fricke) измерил электрическую емкость мембран эритроцитов и получил величину —0,81 мкф/см 2 , что соответствует слою диэлектрика (липида) толщиной 3,3 нм и толщине бимолекулярного слоя, образованного жирными к-тами с 16—17 углеродными атомами.
В 30-х гг. 20 в. Харви и Даниэлли (Е. N. Harvey, J, F, Danielli) показали, что величина поверхностного натяжения на границе жировых капель и цитоплазмы клеток ниже (
0,1 дин/см), чем для чистой границы раздела липид — вода (
10 дин/см). Это указывает на возможность спонтанного образования ассоциатов растворимых белков с ориентированными слоями липидов. Развивая эти представления, Даниэлли и Давсон (H. Davson) в 1935 г. выдвинули первую гипотезу о строении М. б., согласно к-рой мембрана состоит из двойного липидного слоя, покрытого с двух сторон слоями глобулярных белков.
Непосредственная возможность наблюдать биол, мембраны появилась лишь в 50-х гг. 20 в. вследствие развития метода электронной микроскопии и методик приготовления ультратонких срезов. Полученные снимки мембран позволили представить М. б. как трехслойные структуры толщиной порядка 10 нм для плазматических и несколько меньшей — для субклеточных мембран. Робертсон (J. D. Robertson) выдвинул гипотезу об однотипности строения всех биол, мембран и предложил унитарную схему строения мембраны. По Робертсону, белки М- б. могут разворачиваться на поверхности двойного липидного слоя под действием сил электростатического взаимодействия с заряженными головками молекул фосфолипидов; на наружной поверхности мембраны располагаются еще и молекулы гликопротеидов. Эта схема отражала важный принцип строения мембран — ее асимметричность.
Постепенно под влиянием новых фактов, и в первую очередь зернистости структуры мембран, к-рая просматривалась на снимках, полученных при большом увеличении, первоначальные представления о трехслойности мембран были пересмотрены. Вначале Луси (J. Lucy) высказывает предположение о мицеллярной организации липидного слоя в мембране. Шёстрандом (F. S. Sjostrand) была выдвинута гипотеза о глобулярной организации цитоплазматической мембраны в целом. Позже Грин (D. Е. Green) предложил схему организации мембран из субъединиц и сформулировал принцип повторяющихся единиц применительно к внутренней мембране митохондрий. Несмотря на то, что эта модель была тщательно разработана, она не давала удовлетворительного объяснения хорошо известному факту низкой проницаемости мембран для ионов. При этом тетерогенные частицы, образующие мембраны, не удалось получить при фрагментации мембран. Вызывало сомнение и то, что липидам в этой модели отводилась более чем скромная роль, а также практически полное отсутствие учета липид-белкового взаимодействия. В других моделях напр., в модели Бенсона) липид-белковому взаимодействию отводится, напротив, центральная’ роль в формировании мембран.
В дальнейшем оказалось, что отказ от представлений о существовании сплошного бимолекулярного слоя был преждевременным. По новым воззрениям белки не выстилают поверхность липидного слоя, а как бы плавают на поверхности в виде отдельных глобулярных молекул или частиц, в большей или меньшей степени погруженных в мембрану. Эта жидкомозаичная модель, предложенная Ленардом и Сингером (J. Lenard, S. Singer), позволяет удовлетворительно объяснять целый ряд фактов, в частности зависимость многих физиологических функций мембран и активности отдельных мембранных ферментов от фазового состояния липидов в мембране и степени ее текучести (вязкости). Однако белок-белковое взаимодействие в этой модели учитывается недостаточно, и она не позволяет удовлетворительно объяснить экспериментально установленный факт сохранения структуры и основных параметров мембраны при извлечении из нее значительного количества липидов. Эти факты были в дальнейшем учтены в белковокристаллической модели, предложенной Вандеркои (G. Vanderkooi) и Грином, отличающейся фактически лишь наличием протяженных белковых структур, образующихся в результате осуществления дальнодействующих белок-белковых связей.
Наиболее популярными моделями Мембран биологических, стали жидкомозаичные. В то же время становится все более ясным, что мембраны отличаются друг от друга: они разнообразны по составу и специфичны в функциональном отношении. Тонкая организация мембран тесно взаимосвязана с их функциональным состоянием; и то, и другое характеризуется чрезвычайной чувствительностью к действию внешних факторов. При этом нек-рые мембраны проявляют отдельные черты, свойственные различным моделям, а иногда (напр., у нек-рых бактерий) мембраны представляют собой как бы набор фрагментов, соответствующих какой-то одной из разработанных моделей.
Состав и структура биологических мембран
В мембранах, полученных из разных источников, содержание липида колеблется от 25 до 70% (по массе), причем липидный состав многокомпонентен и исключительно изменчив. Единственной общей характеристикой липидов различных мембран является обязательное наличие в их составе так наз. амфипатичных липидов, проявляющих одновременно гидрофильные и гидрофобные свойства. Белковый состав мембран также исключительно разнообразен. Мембраны содержат большое число различных белков с относительным мол. весом (массой) от 25 000 до 230 000. Исключение составляют лишь мембраны палочек сетчатки, содержащие практически один белок — родопсин, и миелин, содержащий три типа белков. В зависимости от степени гидрофобности (т. е. числа и локализации гидрофобных аминокислотных остатков в полипептидной цепи) белки либо частично, либо целиком погружены в липидный слой мембраны и пронизывают его насквозь. В функциональном отношении мембранные белки подразделяют на ферменты, рецепторы, белки транспортных систем и структурные белки. В состав большинства мембран входят также углеводы (до 10% от общей сухой массы мембраны) в форме гликопротеидов и гликолипидов.
Основные компоненты М. б., как правило, синтезируются вне мембранной системы механизмов. Их включение в мембраны еще недостаточно изучено. В наиболее простых случаях, вероятно, участвуют обменные белково-липидные комплексы (напр., фосфолипиды). Возможно и сопряжение процессов биосинтеза с быстрым включением компонентов в мембрану (напр., Na+, К+ — АТФ-азы). По-видимому, отдельные компоненты могут встраиваться в мембраны независимо друг от друга, т. к. в мембранах, как правило, отсутствуют специфические центры роста. Вместе с тем не вызывает сомнений, что нек-рые липид-зависимые ферменты в определенных ситуациях встраиваются в мембрану одновременно с липидным окружением.
Мембранные структуры сформированы за счет сравнительно слабых сил гидрофобных и электростатических (ван-дер-ваальсовых) взаимодействий (см. Молекула, Строение вещества). Ковалентные связи в формировании мембранных структур играют второстепенную роль. В связи с этим мембраны обладают рядом особых физ.-хим. свойств. Так, молекулярные компоненты сохраняют в мембранах довольно высокую подвижность. Различают внутримолекулярную подвижность, связанную с вращательной подвижностью вокруг одиночных связей, вращательную подвижность молекул в целом, движение молекул в плоскости мембраны (латеральная подвижность) и вертикальную подвижность молекулярных компонентов мембран (либо частичную, либо с переходом молекулы из одной половины мембраны в другую).
Обычно мембраны функционируют при температурах, когда жирнокислотные остатки фосфолипидов находятся в жидком (точнее жидкокристаллическом) состоянии (см. Жидкие кристаллы). В этом состоянии скорость диффузии фосфолипидов обеспечивает передвижение липидной молекулы за время порядка одной секунды. Переходы же липидов с одной половины бимолекулярного слоя мембраны на другую (так наз. переходы флип-флоп) совершаются сравнительно редко. Этот процесс, очевидно, должен быть заторможен в мембранах с выраженной асимметрией липидного состава, напр, в мембранах эритроцитов, в к-рых сфингомиелин и фосфатидилхолин находятся главным образом в наружной половине мембраны, а фосфатидилэтаноламин и фосфатидилсерин — на ее внутренней стороне.
Белковые молекулы также проявляют довольно большую свободу движения в мембране. Так, скорость вращательного движения и латеральной подвижности нек-рых гликопротеидов (антигенных и рецепторных белков) соответствует скорости их свободной диффузии в среде соответствующей вязкости. Обнаружена также вертикальная подвижность мембранных белков, обычно сопряженная с их функциональным состоянием. Напр., глубина погружения родопсина в мембрану меняется в зависимости от функционального состояния белка. Свободное движение, однако, присуще не всем мембранным белкам. Часто они образуют устойчивые плотноупакованные образования (область межклеточных контактов, бляшки пурпурных мембран Halobacterium и др.) или строго ориентированные системы (компоненты электрон-транспортных цепей митохондрий, хлоропластов, эндоплазматического ретикулума и др.). Очевидно, подвижность белков ограничена в тех случаях, когда они находятся в липидном микроокружении, отличном по составу от основной массы липидов мембран.
В соответствии с различными типами подвижности мембранных компонентов наблюдается значительная гетерогенность в вязкости различных участков М. б. Низкие значения вязкости наблюдаются в углеводородном слое липидов, причем вязкость убывает к середине слоя в соответствии с градиентом вращательной подвижности углеводородных цепей липидов. Минимальные значения вязкости в середине липидного би-слоя, измеряемой при температурах выше точки фазового перехода, составляют сотые доли пуаз. Вязкость полярного слоя мембраны, измеренная по вращательной и латеральной подвижности молекул, соответствует единицам пуаз. В то же время общая вязкость мембран как структурного образования клетки, измеренная по величине механической деформации мембраны в целом, достигает 10 7 — 10 8 пуаз. Нек-рые другие физ.-хим. свойства мембран приведены в таблице.
Таблица. Сравнение некоторых физико-химических свойств биологических и искусственных фосфолипидных мембран
Биологические мембраны (t° 25°)
Искусственные фосфолипидные мембраны (t° 36°)
Источник