Витамин в2 процессе окисления жирных кислот

CAS номер: 83-88-5
Брутто формула: C17H20N4O6
Внешний вид: кристаллический порошок, темно-желтого цвета, горький на вкус
Химическое название и синонимы: riboflavin,Лактофлавин, Рибофлавин, Витамин B2, 7,8-Dimethyl-10-ribitylisoalloxazine.
Физико-химические свойства:
Температура плавления — 290 гр. Цельсия
Растворимость в воде — 0,07 г / л (20 ºC)
Рибофлавин – водорастворимый витамин, участвующий в своих коферментных формах в окислительно-восстановительных реакциях. Являясь активным центром ферментов – дегидрогеназ и оксидаз, представлен в виде флавинмононуклеотида (ФМН) и флавинадениндинуклеотида (ФАД).
Рибофлавин носит название витамина В2, он необходим для организма и играет роль в выработке энергии, метаболизме жиров и лекарств, а также в нормальном функционировании, росте и развитии клеток. Рибофлавин естественным образом содержится в некоторых пищевых источниках, добавляется к другим продуктам и доступен в качестве пищевой добавки. Рибофлавин также вырабатывается бактериями, присутствующими в толстой кишке во время нормального пищеварения.
Абсорбция рибофлавина происходит преимущественно в проксимальной части тонкой кишки. Недавние исследования показывают, что ФАД и ФМН необходимо преобразовывать в рибофлавин с помощью кистевых пограничных фосфатаз. Абсорбция усиливается с пищей и уменьшается на пустой желудок. Снижение всасывания можно наблюдать при обструктивных заболеваниях желчевыводящих путей, гепатите и циррозе печени. Поглощение рибофлавина в толстой кишке является адаптивным абсорбционным механизмом, зависящим от концентрации в просвете и числа рецепторов в энтероцитах и регулируется энергозависимой транспортной системой.
Дефицит рибофлавина определяется термином как арибофлавиноз. Признаки, связанные с дефицитом, включают хейлоз, угловой стоматит, глоссит, дерматит мошонки и вульвы, себорейный дерматит, кератит и глазные изменения. Тяжелые состояния дефицита связаны с нормохромной, нормоцитарной анемией и невропатией. Клинические признаки и симптомы дефицита развиваются после недостаточного поступления рибофлавина в организм с пищей в течение 3-8 месяцев. Дефицит рибофлавина, отмечаемый во всем мире, распространен в основном в слаборазвитых странах.
В пищевой промышленности рибофлавин используется, как пищевой краситель (Е101), либо в качестве витаминной добавки. Препараты, содержащие рибофлавин, выпускаются в виде капсул, таблеток, в составе мультивитаминных комплексов.
Рибофлавин получают непосредственно из культуры с высоким выходом путем культивирования микроорганизма, продуцирующего рибофлавин, в среде с использованием низшего (С1-С4) алифатического соединения. Затем происходит отделение рибофлавина, содержащегося в культуре, в виде нагретого водного раствора от твердых веществ, и кристаллизация рибофлавина из нагретого водного раствора. Рибофлавин также получают с высоким выходом путем культивирования рибофлавин-продуцирующих дрожжей, принадлежащих к роду Saccharomyces, которые нуждаются в пурине и / или устойчивы к 3-амино-1,2,4-триазолу в среде, и собирают образующийся рибофлавин.
Кроме того, рибофлавин получают с высоким выходом даже в присутствии ионов железа, предварительно выращивая в жидкости дрожжи, продуцирующие рибофлавин, принадлежащие к роду Saccharomyces, и затем выращивая их в среде, продуцирующей рибофлавин, содержащей ионы цинка.
Действие на организм:
В структуре дегидрогеназ участвует в мультиферментной реакции окисления и декарбоксилирования пировиноградной кислоты, как активный центр дегидролипоат-дегидрогеназы возвращается вместе с липоевой кислотой в окисленное состояние в результате чего выделяется НАДН ФАД Участвует в дегидратации янтарной кислоты (сукцината) в составе сукцинатдегдрогеназы и в алфа-кетоглуторатдегидрогеназе для образовании Глутамата из алфа-Кетоглутората в цикле трикарбоновых кислот.
Окисляет жирные кислоты, являясь коферментом ацетил-КоА-дегидрогеназы. Процесс проходит во всех тканях организма, для образования энергии (кроме нервной ткани).
В составе изофермента глицерол-трифосфат-дегидрогеназы участвует в переносе атомов водорода из митохондрий при образовании энергии из глюкозы в печени и скелетных мышцах.
В дыхательной цепи молекулы ФАДН (вместе с НАДН) в процессе окисления передают атомы водорода (ФАД возвращается в окисленную форму) на ферменты дыхательной цепи, которые в дальнейшем переносятся за мембрану митохондрий, расходуя в процессе энергию электронов. Создается разница градиента водорода, которого уже больше за пределами органеллы и уже в реакции фосфорилирования, используя АТФ-синтетазу водород присоединяется к АДФ, образуя АТФ. Таким образом ФАД включается в образование энергии, необходимой для дыхания клетки.
В структуре D-оксидаз кофермент ФАД, а в молекуле L-оксидаз как коэнзим ФМН — приводит к прямому аэробному окислительному дезаминированию аминокислот.
Уничтожает биогенные амины (гистамин, серотонин, дофамин и т.д.) в составе ферментов-оксидаз.
Таким образом, коферментные формы Рибофлавина – ФАД и ФМН являются участниками окислительно-восстановительных реакций, без которых не смогли бы протекать многие важнейшие энергетические процессы в организме.
Гиповитаминоз приводит в первую очередь к поражению высокоаэробных тканей, использующих большое количество энергии – это кожа и слизистые. Вызывается не правильным хранением продуктов, вследствие чего происходит разрушение рибофлавина, алкоголизма, голода и нарушения всасываемости в ЖКТ. Клиническая картина недостатка Витамина В, выражается в виде сухости, воспаления и васкуляризации видимых слизистых оболочек, сухости и шелушения кожи.
Некоторые исследования показали, что добавки рибофлавина могут уменьшить приступы мигрени, и некоторые люди с мигренью принимают riboflavin для ее лечения мигрени, чтобы помочь контролировать возникающие симптомы.
Источник
Для окисления жирных кислот существует свой путь
Окисление жирных кислот (β-окисление)
Для преобразования энергии, заключенной в жирных кислотах, в энергию связей АТФ существует метаболический путь окисления жирных кислот до СО2 и воды, тесно связанный с циклом трикарбоновых кислот и дыхательной цепью. Этот путь называется β-окисление, т.к. происходит окисление 3-го углеродного атома жирной кислоты (β-положение) в карбоксильную группу, одновременно от кислоты отщепляется ацетильная группа, включающая С 1 и С 2 исходной жирной кислоты.
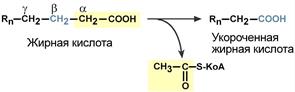
Элементарная схема β-окисления
Реакции β-окисления происходят в митохондриях большинства клеток организма (кроме нервных клеток). Для окисления используются жирные кислоты, поступающие в цитозоль из крови или появляющиеся при липолизе собственных внутриклеточных ТАГ. Суммарное уравнение окисления пальмитиновой кислоты выглядит следующим образом:
Пальмитоил-SКоА + 7ФАД + 7НАД + + 7Н2O + 7HS-KoA → 8Ацетил-SКоА + 7ФАДН2 + 7НАДН
Этапы окисления жирных кислот
1. Прежде, чем проникнуть в матрикс митохондрий и окислиться, жирная кислота должна активироваться в цитозоле. Это осуществляется присоединением к ней коэнзима А с образованием ацил-SКоА. Ацил-SКоА является высокоэнергетическим соединением. Необратимость реакции достигается гидролизом дифосфата на две молекулы фосфорной кислоты.
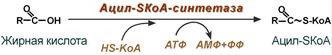
Реакция активации жирной кислоты
2. Ацил-SКоА не способен проходить через митохондриальную мембрану, поэтому существует способ переноса жирной кислоты в комплексе с витаминоподобным веществом карнитином (витамин В11). На наружной мембране митохондрий имеется фермент карнитин-ацилтрансфераза I.
![]()
Карнитин-зависимый транспорт жирных кислот в митохондрию
3. После связывания с карнитином жирная кислота переносится через внутреннюю митохондриальную мембрану транслоказой . На внутренней стороне этой мембраны фермент карнитин-ацилтрансфераза II вновь образует ацил-SКоА, который вступает на путь β-окисления.
4. Процесс собственно β-окисления состоит из 4-х реакций, повторяющихся циклически. В них последовательно происходит окисление (ацил-SКоА-дегидрогеназа), гидратирование (еноил-SКоА-гидратаза) и вновь окисление 3-го атома углерода (гидроксиацил-SКоА-дегидрогеназа). В последней, трансферазной, реакции от жирной кислоты отщепляется ацетил-SКоА . К оставшейся (укороченной на два углерода) жирной кислоте присоединяется HS-КоА, и она возвращается к первой реакции. Все повторяется до тех пор, пока в последнем цикле не образуются два ацетил-SКоА.
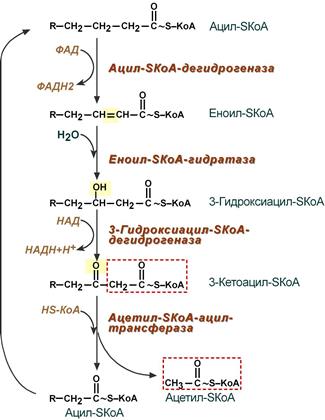
Последовательность реакций β-окисления жирных кислот
Расчет энергетического баланса β-окисления
Ранее при расчете эффективности окисления коэффициент P/O для НАДH принимался равным 3,0, для ФАДH2 – 2,0.
По современным данным значение коэффициента P/O для НАДH соответствует 2,5, для ФАДH2 – 1,5.
При расчете количества АТФ, образуемого при β-окислении жирных кислот необходимо учитывать:
- количество образуемого ацетил-SКоА – определяется обычным делением числа атомов углерода в жирной кислоте на 2.
- число циклов β-окисления . Число циклов β-окисления легко определить исходя из представления о жирной кислоте как о цепочке двухуглеродных звеньев. Число разрывов между звеньями соответствует числу циклов β-окисления. Эту же величину можно подсчитать по формуле (n/2 -1), где n – число атомов углерода в кислоте.
- число двойных связей в жирной кислоте. В первой реакции β-окисления происходит образование двойной связи при участии ФАД. Если двойная связь в жирной кислоте уже имеется, то необходимость в этой реакции отпадает и ФАДН2 не образуется. Количество недополученных ФАДН2 соответствует числу двойных связей. Остальные реакции цикла идут без изменений.
- количество энергии АТФ , потраченной на активацию (всегда соответствует двум макроэргическим связям).
Источник
Параграф 45 -окисление жирных кислот
Автор текстов Анисимова Елена Сергеевна. Авторские права защищены. Продавать текст нельзя.
Курсив не зубрить. Замечания присылать по электронной почте exam_bch@mail.ru
ПАРАГРАФ 45:
«;-ОКИСЛЕНИЕ».
формулы см. в файле «45 ФОРМУЛЫ КРАТКО».
45.0. Определение:
;-окисление – это превращение жирной кислоты в несколько молекул ацетилКоА. Остальное см. далее.
45.1. Непосредственные продукты процесса и их использование.
Этот процесс даёт несколько молекул ацетилКоА, НАДН и ФАДН2. (Это непосредственные продукты ;-окисления).
АцетилКоА после этого используется в ЦТК, что даёт ЭНЕРГИЮ (с помощью дыхательной цепи).
В печени избыток ацетилКоА может использоваться для синтеза кетоновых тел, которые хороши тем, что транспортируются в ткани для выработки энергии, в том числе в головном мозге (при голоде это существенная поддержка для мозга), но опасны при избытке (так как приводят к ацидозу – см. кетозы в п.47).
НАДН и ФАДН2 поступают в дыхательную цепь для выработки энергии в форме АТФ и ТЕПЛА.
45.2. Локализация процесса (в каких клетках и органеллах протекает).
Во всех клетках, кроме эритроцитов и нейронов. В МИТОХОНДРИЯХ (поэтому процесса нет в эритроцитах).
45.3.1. ЗНАЧЕНИЕ ;-окисления жирных кислот для организма –
этот процесс является источником энергии для большинства клеток, в том числе для СЕРДЦА и скелетных мышц (при длительной работе), печени и почек и т.д.. Эритроциты и нейроны не могут использовать в качестве источника энергии процесс ;-окисления жирных кислот. В отличие от глюкозы, способной дать энергию даже при отсутствии кислорода, жирные кислоты дают энергию только в присутствии кислорода, то есть только в аэробных условиях), так как требуется помощь дыхательной цепи.
(не зубрите курсив. Это всё логически выводится)
45.3.2. Последствия недостаточной активности ;-окисления
К каким последствиям приводит недостаточная активность ;-окисления? Последствия недостаточной активности ;-окисления – дефицит энергии, что приводит к мышечной слабости, болях в мышцах.
Особенно страдает СЕРДЦЕ – ему не хватает АТФ для работы (для сокращения мышц), для «восстановительных работ» — синтеза новых молекул взамен повреждённых (окисленных белков и липидов мембран), для работы ионных насосов, что важно для поддержания нормального мембранного потенциала.
Кроме того, при недостаточной активности ;-окисления клетки начинают активнее использовать для выработки энергии глюкозу, что приводит к снижению количества глюкозы в крови (гипогликемии), поэтому глюкозы может не хватать мозгу и эритроцитам (см. гемолиз, п.121 и гипогликемию, п.37).
45.3.3. Причины недостаточной активности ;-окисления:
1) дефицит субстрата – жирных кислот при голоде и низких запасах жира в организме, при истощении,
2) дефицит витаминов (РР, В2, пантотената) и коферментов процесса, снижение активности ферментов из-за мутаций их генов,
3) недостаточная стимуляция процесса гормонами при их дефиците,
4) дефицит КАРНИТИНА (см. далее),
5) повреждения митохондрий активными формами кислорода при дефиците антиоксидантов (см.п.27).
То есть отсутствие или дефицит чего-то, что нужно для ;-окисления.
45.3.4. Коррекция недостаточной активности ;-окисления:
для предотвращения и исправления последствий – соответственно причинам (см. выше): обеспечение организма жирными кислотами за счёт нормализации и питания и процесса расщепления жира (см. регуляцию липолиза), обеспечение витаминами, аминокислотами, карнитином, коррекция дефицита гормонов, защита митохондрий АНТИОКСИДАНТАМИ от повреждения.
45.3.4. Последствия избыточной активности ;-окисления в печени
К каким последствия приводит избыточная активность ;-окисления? Последствия избыточной активности ;-окисления в печени – образование избытка ацетилКоА, который превращается в кетоновые тела, что приводит к ацидозу. При диабете этот ацидоз бывает столь силён, что приводит к коме (если вовремя не принять меры, то есть если не ввести инсулин).
45.3.5. Причины избыточной активности ;-окисления –
недостаточное торможение процесса инсулином при дефиците инсулина при диабете.
Другие причины – чрезмерная стимуляция процесса катехоламинами при стрессе, глюкагоном при голоде, а также катехоламинами, глюкагоном и йодтиронинами (при тиреотоксикозе), ГКС (при кушингизме) при заболеваниях, при которых в организме повышена выработка этих гормонов – см. п.47.
45. 4. Откуда в клетках берутся жирные кислоты для окисления?
Источник жирных кислот для ;-окисления – образование жирных кислот при расщеплении жира (то есть при липолизе). Когда говорят, что жир является энергетическим резервом, имеют в виду, что он может при расщеплении давать жирные кислоты, окисление которых и даёт энергию.
При расщеплении жира в клетках белой жировой ткани жирные кислоты поступают в кровь, с током крови транспортируются к разным клеткам с помощью транспортных белков плазмы крови – АЛЬБУМИНОВ.
Кроме того, жирные кислоты поступают в кровь при расщеплении жира хиеломикронов, которые присутствуют в крови после приёма жирной пищи. См. п.49-51.
При расщеплении жира в клетках бурого жира жирные кислоты не поступают в кровь, а окисляются в тех же клетках ради образования энергии в форме тепла (благодаря разобщению окислительного фосфорилирования – см. п.27 и 44).
45.5. Регуляция ;-окисления жирных кислот гормонами. И её значение.
Регуляция гормонами.
;-окисление активируется при СТРЕССЕ под влиянием катехоламинов адреналина и норадреналина и при ГОЛОДЕ под влиянием гормона голода глюкагона, чтобы дать энергию (АТФ) в этих состояниях.
Кроме того, стимуляция процесса этими гормонами приводит к образованию в печени избытка ацетилКоА, что приводит к синтезу кетоновых тех и создаёт риск кетоза. Поэтому избыток гормонов, стимулирующих ;-окисление, создаёт опасность кетоза.
В состоянии сытости ;-окисление жирных кислот снижается под влиянием ИНСУЛИНА (гормона покоя и сытости).
Замедляя ;-окисление, инсулин предотвращает накопление кетоновых тел (кетоз) – поэтому при снижении количества инсулина при диабете первого типа риск кетоза повышается – из-за активации ;-окисления на фоне снижения тормозящего влияния инсулина. При кетоацидотической коме главным лекарством является инсулин – см. п.103.
45.6. Реакции ;-окисления жирных кислот. См. формулы в «45 формулы».
1. Процесс происходит в митохондриях, поэтому сначала жирную кислоту нужно перенести из гиалоплазмы в митохондрию.
Перед переносом жирной кислоты в митохондрию её нужно АКТИВИРОВАТЬ.
Активация (активирование) жирных кислот заключается в их реакции с коферментом А (переносчиком ацилов). СМ. РЕАКЦИИ В ТАБЛИЦЕ К ПАРАГРАФУ 45 – только так, «с картинками», есть смысл читать описание реакций.
При реакции с коферментом А атом углерода карбоксильной группы соединяется с атомом серы кофермента А сложноэфирной связью, при этом атом серы присоединяется «вместо» ОН атомов карбоксильной группы. Образуется соединение остатка жирной кислоты (то есть части молекулы жирной кислоты без ОН атомов карбоксильной группы, которая называется ацилом) с коферментом А, которое называется – ациломКоА. (Не путать с ацетиломКоА – у ацетила 2 атома углерода, а ацил – это более широкое понятие – ацилами называются остатки карбоновых кислот с любым количеством атомов углерода – хоть 4, хоть 16).
Образование связи между ацилом и коферментом А происходит с затратой энергии, которая выделяется при расщеплении АТФ до АМФ и ФФн (то есть при отщеплении от АТФ двух фосфатов). В таких случаях затраты АТФ считают за ДВЕ молекулы АТФ. Так как АМФ вступает в реакцию с АТФ, которая приводит к образованию 2 молекул АТФ – в этой реакции и происходит превращение второй молекулы АТФ в АДФ).
Фермент. Ферменты, которые катализируют синтезы с затратой АТФ, называются синТЕтазами. Синтетаза, которая катализирует синтез ацилКоА, называется синтетазой ацилаКоА или ацилКоА/синтетазой.
2. Ацил переносится с кофермента А на КАРНИТИН, который осуществляет перенос ацила через мембрану митохондрии:
ацилКоА + карнитин ; КоА + ацил/карнитин.
На внутренней стороне мембраны митохондрии ацил переносится на молекулу кофермента А, которая находится внутри митохондрии:
Ацил/карнитин + кофермент А ; ацилКоА + карнитин.
После этого ацилКоА, находящийся в митохондрии, вступает в реакции окисления.
(реакции 3.1-3.3 – пример процесса так называемого КОСВЕННОГО КИСЛЕНИЯ – то есть «окисления кислородом воды»;
другие примеры косвенного окисления – превращение сукцината к оксалоацетат в ЦТК и превращение этанола в уксусную кислоту)
3.1. От ацилКоА отщепляются 2 атома водорода (от 2-го и от 3-го атомов углерода), в результате чего образуется соединение с двойной связью между 2-м и 3-м атомом углерода, которое называется еноиломКоА. («ен» означает двойную связь).
Отщеплённые 2 атома водорода переносятся на кофактор ФАД (содержащий витамин В2), в результате чего ФАД превращается в ФАДН2. От ФАДН2 атомы водорода передаются по дыхательной цепи на кислород. В результате из водорода и кислорода образуется одна молекула воды. А за счёт энергии, выделяющееся при работе дыхательной цепи (при переносе электронов) образуются 1,5 молекулы АТФ.
Если кислорода нет, если В2 нет, то ;-окисление протекать не может.
Процесс отщепления водорода называется дегидрированием.
Ферменты, катализирующие отщепление атомов водорода от субстрата и их перенос на НАД+ или ФАД (или НАДФ+), называются дегидрогеназами. Дегидрогеназа, отщепляющая водород от ацилаКоА, называется дегидрогеназой ацилаКоА или ацилКоА/дегидрогеназой.
3.2. ЕноилКоА вступает в реакцию с молекулой воды (НОН);
при этом ОН присоединяется к 3-му атому углерода еноилаКоА, а атом Н воды – ко 2-му атому углерода еноилаКоА («по» двойной связи).
В результате вещество с ОН (гидрокси) группой в 3-м (;) положении, которое называется ;-гидрокси/ациломКоА.
Реакция присоединения НОН «по» двойной связи называется гидратацией, а катализрующий её фермент называется гидратазой. Гидратаза еноилаКоА называется еноилКоА/гидратазой.
3.3. От ;-ГИДРОКСИацилКоА отщепляются 2 атома водорода (от кислорода ОН группы и от 3-го атома углерода), в результате чего образуется соединение с двойной связью между атомом кислорода и 3-м атомом углерода (С=О), то есть с КЕТОгруппой, которое называется ;-КЕТОациломКоА.
Отщеплённые 2 атома водорода переносятся на кофактор НАД+ (содержащий витамин рр), в результате чего НАД превращается в НАДН,Н+. От НАДН,Н+ атомы водорода передаются по дыхательной цепи на кислород. В результате из водорода и кислорода образуется одна молекула воды. А за счёт энергии, выделяющееся при работе дыхательной цепи (при переносе электронов) образуются 2,5 молекулы АТФ.
Если кислорода нет, если РР нет, то ;-окисление протекать не может.
Процесс отщепления водорода называется дегидрированием.
Ферменты, катализирующие отщепление атомов водорода от субстрата и их перенос на НАД+ или ФАД (или НАДФ+), называются дегидрогеназами. Дегидрогеназа, отщепляющая водород от ;-гидрокси/ацилаКоА, называется дегидрогеназой ;-гидрокси/ацилаКоА или ;-гидрокси/ацилКоА/дегидрогеназой.
Таким образом, превращение ацилКоА в кетоацилКоА даёт 4 молекулы АТФ – за счёт реакций 3.1 и 3.3.
3.4. В молекуле ;-КЕТОацилаКоА происходит расщепление связи между 2-м и 3-м атомами углерода,
в результате молекула расщепляется на два фрагмента – двухуглеродный и 14-тиуглеродный (если рассматривать окисление 16-ти углеродной пальмитиновой кислоты).
Это расщепление молекулы происходит при её реакции с молекулой кофермента А (КоА-SH).
К двухуглеродному отщеплённому фрагменту присоединяется атом водорода (ко второму атому углерода, «по месту расщеплённой связи»), в результате чего фрагмент превращается в молекулу ацетилКоА. Это ПЕРВАЯ из 8 молекул ацетилКоА, которые образуются при ;-окислении пальмитата.
Ко второму фрагменту присоединяется атом серы кофермента А (к бывшему 3-му атому углерода расщеплённой молекулы),
в результате чего фрагмент превращается в 14-углеродную молекулу ацилКоА.
4. 14-углеродный ацилКоА подвергается тем же реакциям, которым подвергся 16-углеродный ацилКоА (то есть реакциям 3.1-3.4), в результате чего образуются: ВТОРАЯ молекула ацетилКоА и 12-углеродная молекула ацилКоА. А также 4 молекулы АТФ за счёт двух коферментов.
12-углеродная молекула ацилКоА в свою очередь тоже подвергается реакциям, аналогичным 3.1-3.4, в результате чего образуются ТРЕТЬЯ молекула ацетилКоА и 10-углеродная молекула ацетилКоА, а также 4 молекулы АТФ.
И т.д. – всё это происходит до тех пор, пока не получится 4-углеродная молекула ацилКоА, которая после реакций 3.1-3.4 даёт 2 молекулы ацетилКоА (отщепляется СЕДЬМАЯ молекула ацетилКоА и остаётся молекула ацетилКоА, которая является восьмой).
45.7. Подсчёт АТФ от ;-окисления одного пальмитата.
Таким образом, при окислении одной молекулы пальмитиновой кислоты реакции 3.1-3.4 происходят СЕМЬ раз –
семь раз отщепляется молекула ацетилКоА,
7 раз образуется по 4 молекул АТФ (за счёт 7 молекул ФАДН2 и 7 молекул НАДН, Н+, с помощью дыхательной цепи, с использование кислорода), то есть всего образуется 28 молекул АТФ.
8 молекул ацетилКоА поступают в ЦТК, что приводит (с помощью дыхательной цепи) к образованию ещё 10 молекул АТФ от каждой молекулы ацетилКоА, то есть 8*10=80 молекул АТФ.
Если витаминов ЦТК и ДЦ мало (РР, В2, пальмитата, В1, липоевой кислоты), то энергии мало.
Таким образом, всего при окислении пальмитата образуется 28+80=108 молекул АТФ.
Нужно вычесть из 108 молекул АТФ те 2 молекулы АТФ, которые были затрачены на активацию жирной кислоты.
В итоге получается, что одна молекула пальмитата даёт 108 – 2=106 молекул АТФ.
45.8. Подсчёт АТФ от использования одной молекулы жира.
Молекула жира при расщеплении (липолизе – п.44) даёт одну молекулу глицерина и 3 молекулы жирных кислот.
Поэтому жир даёт 106*3 молекул АТФ = 318 АТФ.
Плюс АТФ от окисления глицерина (17,5). В итоге более 300 (335,5) молекул АТФ от одной молекулы жира – это в 10 раз больше, чем от полного аэробного окисления одной молекулы глюкозы (32 АТФ – п.32).
45.9. Подсчёт АТФ от ;-окисления стеариновой кислоты.
При ;-окислении не пальмитиновой, а стеариновой кислоты происходят аналогичные реакции,
но первый ацил содержит не 16, а 18 атомов углерода, поэтому даёт не 8, а 9 молекул ацетилКоА;
не 7, а 8 раз по 4 АТФ от коферментов и т.д. – распишите это самостоятельно (9ацетилКоА по 10 АТФ – это 90, 8 коферментов 4 АТФ – это 32 АТФ, 90+32=122, минус 2 АТФ на активацию — 122-2=120 АТФ от одной молекулы стеарата).
45.10. Как запомнить реакции процесса 45 и 46.
1. Выучить формулу жирной кислоты: СН3 – (СН2 )14 – СООН. Или не 14, а 16.
2. Выучить формулу эфира жирной кислоты с коферментом А, то есть ацилКоА:
СН3 – (СН2 )14 – С(О)SКоА.
3. Запомнить последовательность метаболитов: ацилКоА, еноилКоА, ГИДРОКСИацилКоА и КЕТОацилКОА.
4. Знать, что ен – это двойная связь, что гидрокси – это ОН группа, что кето – это С=О группа.
Научиться рисовать ацил с этими группами.
Помнить, что ;- это ТРЕТИЙ атом углерода, а не второй.
И что процесс называется ;-окислением именно потому, что окисление происходит «по» бета-положению, то есть по 3-му атому углерода.
5. Найти отличия между формулами начального и конечного метаболита реакции и на основе этого отличия догадаться, какие атомы присоединились к веществу или отсоединились от вещества. Как в детской игре «найди 2 отличия между похожими картинками».
6. Помнить, что если отщепились атомы водорода, то они переносятся на кофермент (см. 3.1 и 3.3).
7. Помнить, что если атомы водорода присоединились к метаболиту, то скорее всего они перенесены на него от кофермента (см. п.46).
Источник