Витамин в1 механизм всасывания
Всасывание макромолекул. Крупные молекулы и их агрегаты всасываются в кишечнике по механизму трансцитоза. В энтероцит они поступают путем эндоцитоза. В везикуле, образовавшейся из участка мембраны клетки, вещество транспортируется через цитоплазму энтероцита и выделяется из него в межклеточное пространство путем экзоцитоза. При этом мембрана везикулы (вакуоли), содержащей макромолекулы транспортируемых веществ, «встраивается» в мембрану энтероцита. Сокращения ворсинок способствуют эндоцитозу. Так в кишечнике транспортируются иммуноглобулины, витамины, ферменты, а у новорожденных — белки грудного молока.
Всасывание микромолекул энтероцитами осуществляется по механизму пассивного транспорта (диффузии и осмоса, облегченной диффузии и фильтрации), а также активного транспорта.
Движение молекул через полупроницаемые биологические мембраны энтероцитов в процессе диффузии и осмоса происходит вследствие концентрационных градиентов веществ в цитоплазме клетки и внеклеточной среде. Для облегченной диффузии необходимо наличие мембранных переносчиков.
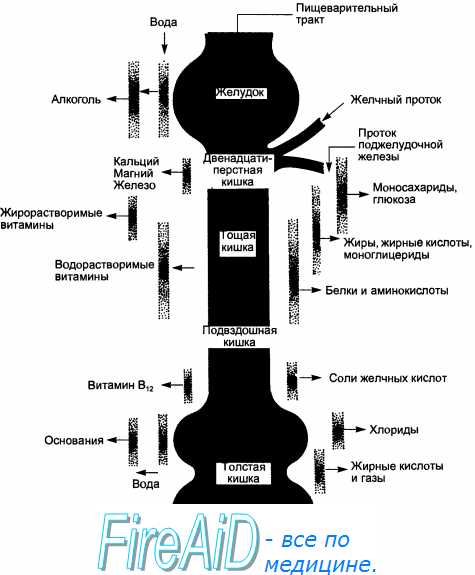 Рис. 11.5. Топография всасывания различных веществ в разных последовательных отделах пищеварительного тракта. Стрелки указывают, что перечисленные вещества поступают из полости пищеварительного тракта во внутреннюю среду организма.
Рис. 11.5. Топография всасывания различных веществ в разных последовательных отделах пищеварительного тракта. Стрелки указывают, что перечисленные вещества поступают из полости пищеварительного тракта во внутреннюю среду организма.
Процесс фильтрации растворенных в воде веществ осуществляется в силу разности давления жидкости над мембраной, выполняющей роль фильтра, и под ней. О существовании механизма фильтрации при всасывании веществ в кишечнике свидетельствует увеличение скорости всасывания изотонического раствора натрия хлорида при повышении внутрики-шечного давления в процессе осуществления моторики кишки.
Двигательная активность кишечника способствует всасыванию продуктов гидролиза пищевых веществ еще и потому, что она обеспечивает перемешивание пристеночного слоя содержимого кишечника.
Процесс всасывания регулируется с помощью нервных и гуморальных механизмов. При механическом раздражении ворсинок тонкой кишки и под влиянием продуктов гидролиза пищевых веществ (пептидов, аминокислот, глюкозы) они резко усиливают и учащают свои сокращения. Этот эффект сохраняется и после перерезки парасимпатических и симпатических нервов кишечника в опытах на животных, что говорит о его реализации через энтеральную нервную систему.
Редкая и слабая сократительная активность ворсинок кишки голодного животного значительно оживляется при переливании ему крови сытого животного. Это свидетельствует о наличии гуморальных регуляторов всасывания. Установлено, что мощным гуморальным стимулятором активности ворсинок является гормон вилликин, выделяемый эндокринными клетками слизистой оболочки двенадцатиперстной кишки.
Обобщенная схема, отражающая топографию всасывания различных веществ, представлена на рис. 11.5.
Из данного рисунка следует, что зоны всасывания различных веществ частично или полностью перекрываются. В желудке всасываются в основном вода, соединения меди, алкоголь. Продукты гидролиза белков, жиров и углеводов всасываются в тех же отделах пищеварительного тракта, где осуществляется их переваривание, т. е. в двенадцатиперстной и тощей кишке. Жирорастворимые и водорастворимые витамины (кроме витамина В12) всасываются в тощей кишке. В подвздошной кишке осуществляется всасывание солей желчных кислот и витамина В12. Двухзарядные катионы (цинка, кальция, магния и железа) всасываются в основном в двенадцатиперстной кишке, а вода, хлориды, основания, жирные кислоты и газы — в толстой кишке.
Источник
Витамин В1
Опубликовано чт, 20/06/2019 — 14:56

Тиамин (витамин B1) является важным питательным веществом, которое служит кофактором для ряда ферментов, в основном с митохондриальной локализацией. Некоторые зависимые от тиамина ферменты участвуют в энергетическом метаболизме и биосинтезе нуклеиновых кислот, тогда как другие являются частью антиоксидантного механизма. Мозг очень уязвим для дефицита тиамина из-за его зависимости от митохондриальной продукции АТФ. Это положение более очевидно во время быстрого роста детей, при котором дефицит тиамина обычно связан с недоеданием или генетическими дефектами. Дефицит тиамина способствует возникновению ряда расстройств неврологических и психопатологических симптомов (спутанность сознания, снижение памяти и нарушения сна) до тяжелой энцефалопатии, атаксии, застойной сердечной недостаточности, мышечной атрофии и даже смерти.
Основным питательным веществом тиамина (витамин B1) является водорастворимый серосодержащий витамин, принадлежащий к комплексу витаминов группы B. Не будучи эндогенно синтезированным, единственным доступным источником тиамина являются некоторые продукты питания (говядина, птица, крупы, орехи и бобы). Организм не хранит тиамин > 30 мг, а период полураспада для тиамина составляет всего 9–18 дней. При среднем потреблении продуктов питания на уровне в 2000 ккал,( потребляемом ежедневно), минимальная потребность в тиамине составляет 0,66 мг , однако, рекомендуемая суточная доза для взрослых мужчин и женщин составляет 1,2 и 1,1 мг соответственно. Во время беременности или кормления грудью потребность в витамине В1 увеличивается до 1,4 мг / день. У детей рекомендуемая диета (RDA) зависит от возраста и составляет от 0,2 мг (от рождения до 6 месяцев) до 0,6 мг (от 6 месяцев до 8 лет). В организме человека богатыми тиамином ткани являются скелетные мышцы, сердце, печень, почки и мозг.
В развитых странах преобладающее использование промышленной обработки пищевых продуктов часто истощает содержание тиамина наряду с другими витаминами и питательными веществами. Повышенное потребление обработанных пищевых продуктов в форме простых углеводов, не дополненных адекватными уровнями тиамина, было названо «калорийным недоеданием». С другой стороны , по меньшей мере, у 29% пациентов с ожирением, которым предстоит хирургическое операции на бариатрической стадии, отмечается как дефицит тиамина. Поскольку тиамин является ключевым фактором метаболизма глюкозы, увеличение потребления углеводов будет пропорционально увеличивать диетическую потребностьи в тиамине (минимум 0,33 мг на 1000 ккал). Таким образом, вместо того, чтобы сосредоточиться на RDA тиамина, важно сопоставить его потребление с потреблением углеводов, а также с общим потреблением калорий.
В развивающихся странах дефицит тиамина остается широко распространенной проблемой из-за высоких показателей потребления белого риса. Поскольку домашние методы измельчения заменяются промышленным измельчением и переработкой риса, важные питательные вещества (такие как тиамин) в отрубях удаляются. Азиатские страны потребляют около 90% риса, произведенного во всем мире, реализуя , по оценкам специалистов , 60% суточной потребности населения в потреблении энергии с пищей, и, следовательно, дефицит тиамина достаточно распространен среди 15% подросткового населения. Дефицит тиамина может развиться при употреблении в пищу диет, загрязненных метаболизирующими тиамин ферментами (например, тиаминазой) или подверглись инактивации тиамина нагреванием и / или диоксидом серы. Чрезмерное употребление танинсодержащих или пищевых продуктов, богатых кофеином, теобромином и теофиллином (например, содержащихся в кофе, шоколаде и чае соответственно), может инактивировать тиамин, тем самым нарушая его статус.
Другие факторы риска , которые увеличивают вероятность недостаточного потребления тиамина включают старение, низкий экономический статус, расстройства пищевого поведения, соматические заболевания , в частности, влияющие на желудочно — кишечный тракт, искусственное питание, бариатрическая хирургия, диабет и злоупотребление алкоголем. Сообщается о неудовлетворенных потребностях в увеличении потребления тиамина в пищу во время лактации, беременности и повышенной физической активности. Во время лактации у детей повышается риск развития авитаминоза , в частности, у дефиците тиамина у матерей. Например, у 27% женщин детородного возраста в Камбодже имеет место дефицит тиамина, а у 38% младенцев диагностирован дефицит тиамина, что является критической проблемой, которая в значительной степени способствует смертности 3-месячных детей. Однако даже при наличии достаточного потребления тиамина его дефицит может быть обусловлен генетическими факторами, то есть патогенными мутациями генов в ключевых регуляторах пути трансформации тиамина, включая тиаминпирофосфокиназу 1 (ТПК1), тиаминдифосфаткиназу (TDPK), тиаминтрифосфатазу (THTPA) и переносчики тиамина (SLC25A19, SLC19A2 / THTR1 и SLC19A3 / THTR. также было показано, что переносчик органических катионов 1 (OCT1) действует как транспортер тиамина в печени.
Независимо от основной причины дефицита тиамина большинство симптомов проявляются на неврологическом уровне. Дефицит тиамина может вызвать повреждение тканей головного мозга, ингибируя использование энергии мозга, учитывая критическую роль тиамин-зависимых ферментов, связанных с использованием глюкозы. Это подтверждается значительным уровнем поглощения тиамина гематоэнцефалическим барьером, что подчеркивает высокую потребность мозга в тиамине и потребность в его снабжении для поддержания адекватных функций мозга, особенно в тех областях мозга, где требуются высокий уровень метаболизма и оборот тиамина.
Как и у большинства гидрофильных микроэлементов, поглощение тиамина происходит в основном в тонком кишечнике. В пищеварительном тракте пищевые белки гидролизуются, выделяя тиамин. В просвете кишечника щелочные фосфатазы катализируют гидролиз тиаминфосфорилированных производных в свободный тиамин. Нефосфорилированный свободный тиамин в концентрациях, превышающих 1 мкМ, поступает в энтероцит путем пассивной диффузии, тогда как на более низких уровнях он транспортируется через систему насыщения тиамином / H + ( переносчик тиамина 1 или THTR1) в зависимости от количества энергии. В условиях дефицита тиамина в клетках Caco2 в культуре наблюдалась активация экспрессии переносчика тиамина 2 (THTR2), что позволяет предположить, что диета может модулировать экспрессию этого транспортера . Внутри энтероцита тиамин фосфорилируется до тиаминпирофосфата (TPP) с помощью TPK1. Затем большая часть TPP дефосфорилируется до тиаминмонофосфата (TMP), чтобы пересечь базальную мембрану энтероцита. TMP высвобождается в кровоток через ATPase-зависимую транспортную систему. Свободный тиамин также может попасть в кровь через транспортер тиамина 2 (THTR2), расположенный в основном на базолатеральной мембране энтероцита. Попадая в кровь, в то время как очень низкие уровни TMP и тиамина циркулируют свободно в плазме или сыворотке, более 90% фосфорилированного тиамина (в форме TPP) присутствует в эритроцитах и лейкоцитах. Примечательно, что изоформа 3 носителя SLC44A4 недавно была описана как носитель TPP в толстой кишке. Первоначально SLC44A4 был описан как транспортер холина, связанный с не нейрональным синтезом холина и необходимый для эфферентной иннервации волосковых клеток в оливо-кохлеарном пучке для поддержания физиологической функции наружных волосковых клеток и защиты волосковых клеток. от акустической травмы. Последние данные указывают на то, что этот носитель может опосредовать абсорбцию микробиоты, генерируемой TPP (особенно у младенцев), и способствовать гомеостазу тиамина хозяина.
Клеточное поглощение тиамина из кровотока может быть опосредовано любым из двух высокоаффинных носителей: THTR1 (кодируется SLC19A2) и THTR2 (кодируется SLC19A3). Эти транспортеры выражены повсеместно, но THTR1 наиболее распространен в кишечнике, скелетных мышцах, нервной системе и глазах, за которым следуют плацента, печень и почка, тогда как THTR2 находится в основном в жировой ткани, печени, лимфоцитах, селезенке, желчном пузыре, плаценте. поджелудочной железе и мозге. После внутриклеточной транспортировки свободный тиамин быстро фосфорилируется до TPP с помощью тиаминпирофосфокиназы (TPK1). Вторая киназа, TDPK, добавляет фосфатную группу к TPP для генерирования тиаминтрифосфата (TTP). TPP и TTP могут быть дефосфорилированы, соответственно, до TMP и TPP с помощью фосфатаз — простатической кислой фосфатазы (ACPP) и THTPA соответственно.
До 90% от общего количества тиамина в организме остается в его дифосфате , метаболически активной форме (TPP), тогда как остальное находится в виде TMP и TTP. TPP является кофактором нескольких тиамин-зависимых ферментов, участвующих в метаболизме углеводов и жирных кислот, а именно цитозольной транскетолазы (TKT), пероксисомальной 2-гидроксиацил-CoA лиазы 1 и трех митохондриальных ферментов (пируватдегидрогеназы, α-кетоглутаратдегидрогеназы и разветвленных). -цепные α-кетокислотные дегидрогеназные комплексы. Биохимическая роль TPP хорошо понятна, но биологическая значимость и вклад TTP не совсем ясны. Ранее считалось, что это специфическая нейроактивная форма тиамина, но в последнее время сообщалось, что TTP (составляет ∼10% от общего пула тиамина мозга) участвует в возбудимости мембраны и нервной проводимости, действуя в качестве модулятора проницаемости хлоридно-натриевых каналов.
В цитозоле TPP действует как кофактор для TKT, ключевого фермента неокислительной ветви пентозофосфатного пути (PPP). Этот метаболический путь генерирует никотинамидадениндинуклеотидфосфат (NADPH) и рибозо-5-фосфат (R5P). NADPH является ключевым восстановителем в биосинтетических реакциях и является одним из субстратов биосинтетических ферментов (синтез жирных кислот) и антиоксидантных ферментов, таких как глутатионпероксидаза-редуктазная система и тиоредоксинпероксидазы.Важное участие R5P в биосинтезе ДНК и РНК подчеркивает критическую роль тиамина в высокопролиферирующих тканях.
Исходя из его роли в биохимических путях, предполагается , что дефицит тиамина приведет к усилению окислительного стресса и снижению пролиферации клеток, а также к снижению синтеза жирных кислот (включая миелин) с тяжелыми последствиями, особенно во время развития мозга. В соответствии с этим предположением, дефицит тиамина снижает активность TKT и приводит к нарушению PPP и снижению нейрогенеза в коре и гиппокампе во время развития нервной системы.
Пероксисомы играют важную роль в катаболизме перекиси водорода, а также в укорочении очень длинных жирных кислот (которые не могут подвергаться прямому катаболизму β-окисления митохондрий) и α-окислении. В последнем процессе TPP-зависимый фермент 2-гидроксиацил-СоА лиаза 1 (HACL1) катализирует расщепление 3-метилразветвленных и 2-гидрокси длинноцепочечных жирных кислот с прямой цепью. Фитановая кислота (3-метилзамещенная 20-углеродная жирная кислота с разветвленной цепью), в отличие от большинства жирных кислот, не может подвергаться β-окислению из-за наличия метильной группы в положении 3. Как таковой, он расщепляется HACL1 в результате первоначального α-окисления. Эта жирная кислота с разветвленной цепью получается из рациона, особенно из молочных продуктов и красного мяса. Нарушение катаболизма фитановой кислоты из-за неадекватных уровней TPP приводит к накоплению триглицеридов, что может вызывать негативные эффекты, такие как мозжечковая атаксия, периферическая полиневропатия, потеря зрения и слуха, аносмия, а в некоторых случаях дисфункция сердца и эпифизарная дисплазия. Симптомы, вызванные дефицитом тиамина, характерны для болезни Рефсума, которая вызвана патогенными мутациями в HACL1. Некоторые из симптомов также наблюдаются при аутосомно-рецессивном системном расстройстве, синдроме Зеллвегера и других пероксисомальных заболеваниях, включая неонатальную адренолейкодистрофию. Синдром Зеллвегера вызван патогенными мутациями в генах пексинов, которые кодируют белки, необходимые для сборки функциональных пероксисом. Он характеризуется дефицитом пути окисления пероксисомных жирных кислот, вызывающего тяжелую неврологическую и печеночную дисфункцию, а также черепно-лицевые нарушения.
Большая часть (∼90%) цитозольного TPP транспортируется в митохондрии с помощью митохондриального переносчика тиаминпирофосфата MTPPT, продукт гена SLC25A19. Этот транспортер обеспечивает обмен цитозольного TPP на митохондриальный TMP; Попав в цитозоль, TMP метаболизируется и превращается обратно в TPP. В митохондриях TPP является критическим кофактором для трех ферментов, а именно пируватдегидрогеназы, α-кетоглутаратдегидрогеназы и α-кетокислотной дегидрогеназы с разветвленной цепью (PDH, αKGDH и BCKDH, соответственно).
Пируватдегидрогеназный комплекс — мультисубъединичный комплекс катализирует TPP- зависимое декарбоксилирование пирувата, генерируя ацетил-КоА, который затем входит в цикл Кребса. Регуляция активности PDH представляет собой ключевой метаболический «переключатель», влияющий на выбор «топлива», то есть между окислением жирных кислот и гликолитическим потоком. Было высказано предположение, что неспособность регулировать выбор топлива для производства метаболической энергии лежит в основе «метаболической жесткости», приводящей к метаболическим нарушениям. Следовательно, опосредованное тиамином ингибирование комплекса PDH блокирует систему в окислении глюкозы в пируват, что приводит к увеличению лактата и снижению клеточной продукции АТФ. Как и ожидалось, в тяжелых случаях метаболический дефицит проявляется как фатальный лактоацидоз у новорожденных, тогда как в более легких случаях неврологические состояния могут приводить к структурным нарушениям в центральной нервной системе (ЦНС), судорогам, умственной отсталости и спастичности.
В случае дефицита тиамина наиболее пораженными участками мозга, по-видимому, являются мозжечок, мамиллярные тела, таламус, гипоталамус и ствол мозга у взрослых. Что касается дефицита тиамина, Zhao et al.(2009) показали, что у мышей депривация тиамина в течение 14 дней приводила к различной степени дефицита ферментов при тестировании на активность TKT, PDH и αKGDH в коре и гиппокампе.
Патогенные мутации в генах, кодирующих ферменты и транспортеры, участвующие в метаболизме тиамина, приводят к симптомам, сходным с теми, что обнаруживаются при дефиците тиамина на основе питания, и перекрываются с нарушениями митохондриальной дисфункции . Эти мутации, затрагивающие гены, ответственные за транспортеры тиамина 1 ( SLC19A2 ; OMIM 249270) и 2 ( SLC19A3 ; OMIM 607483), составляют основную причину субоптимального всасывания тиамина в кишечнике и, как следствие, недостаточного клеточного распределения тиамина по организму.
Как указано выше, патология дефицита тиамина влечет за собой нарушение выработки энергии из митохондрий в форме АТФ при использовании субстратов, генерирующих пируват (например, глюкозы), а также повышенный окислительный стресс. В этих условиях глюкоза черезгликолиз образует пируват, который не может войти в цикл Кребса в виде ацетил-КоА из-за низкой активности PDH. Как таковой, пируват трансаминируется в Ala или восстанавливается до лактата спомощью лактатдегидрогеназы. Это согласуется с повышенным уровнем лактата и органических кислот, наблюдаемых в CSF, моче и крови при дефиците тиамина.
Центральная нервная система человека обладает высокой потребностью в энергии: 2% массы тела контролируют около 20% общих метаболических расходов, большая часть которых расходуется на потенциалы возбуждающего действия, на передачу сигналов между нейронами, через химические синапсы, рост аксонов. и миелинизацию. Поскольку глюкоза является основным «топливом» для производства энергии в головном мозге, неудивительно, что митохондриальная дисфункция и последующее нарушение метаболизма глюкозы связаны с несколькими неврологическими расстройствами и нарушениями развития нервной системы и основными психическими заболеваниями, такими как депрессия и шизофрения.
Неврологические симптомы при дефиците тиамина сходны с дефектами, которые чаще всего проявляются как синдром Ли-Ли с вовлечением базальных ганглиев. Следовательно, нервная система, которая специализируется на использовании глюкозы для выработки энергии, кажется наиболее уязвимой для дефицита PDHC из-за истощения TPP. В мозге плохое производство АТФ в митохондриях будет ограничивать поддержание мембранного потенциала посредством действия Na + , K +-АТФазы, тем самым нарушая нервную проводимость и процессы в синапсах. Кроме того, повышенный окислительный стресс из-за более низкой активности TKT повредит критические биомолекулы, инициируя перекисное окисление липидов и окислительное повреждение белков, что приводит к фрагментации, посттрансляционным модификациям и перекрестным связям. Модификация эпитопов на нормальных, эндогенных молекулах может приводить к активации микроглии и иммунных клеток, усугубляя вызванное окислительным стрессом повреждение.
Уровни тиамина в крови и CSF предоставляют ограниченную информацию при оценке состояния тиамина у субъекта, поскольку они не обязательно отражают метаболическую функцию тиамина или прямую связь с его уровнями в тканях. Таким образом, оценки TKT эритроцитов и, если возможно, оценки других тканеспецифичных TPP-зависимых ферментов (PDH, αKGDH) считаются золотыми стандартами. Базовая активность TKT обычно выражается в единицах на грамм гемоглобина (г Hb), но, что более важно, рассчитывается процент активации TKT в добавках к TPP (0-15% считаются нормальными).
Коэффициент активации TKT (эритроциты) и / или активность TPP-зависимых ферментов (лейкоциты, фибробласты кожи и биопсия мышц) обычно сопровождаются тестированием уровней лактата и пирувата в сыворотке, BCAA, органических кислот, а также методами визуализации мозга. Единственными случаями, когда оценка свободного тиамина в плазме / сыворотке и CSF, по-видимому, является ценным диагностическим инструментом, являются случаи патогенных мутаций в SLC19A3. Точно так же экскреция тиамина с мочой также не является надежным методом для оценки его уровня в организме, поскольку он зависит от его потребления и всасывания. Как правило, он выражается в расчете на единицу креатинина для учета функции почек, и следует учитывать возраст, так как нормальные значения у детей различаются [120 нмоль / ммоль креатинина в возрасте 1–13 лет] и взрослые [220 нмоль / ммоль креатинина в возрасте> 18 лет ].
К сожалению, ранние симптомы дефицита тиамина не выражены или недостаточно различимы, чтобы поставить прямой диагноз. Они включают потерю аппетита, тошноту, слабость, апатию, усталость, раздражение, нарушения сна, анорексию и дискомфорт в животе. Кроме того, выявление конкретных клинических симптомов дефицита тиамина является проблематичным, поскольку оно скрывается за счет влияния других сопутствующих состояний (сопутствующих заболеваний), таких как инфекции и / или разнообразные нарушения питания.
Клиническая классификация дефицита тиамина обычно делится на «сухую» (или невритическую, характеризующуюся полиневропатией, сниженным коленным рефлексом и другими сухожильными рефлексами и прогрессирующей сильной слабостью мышц) и «влажную» (или сердечную, характеризующуюся отеком ног, тела и лица высокий сердечный выброс, желудочковая недостаточность и застой в легких).
При раннем подозрении на генерализованный дефицит тиамина рекомендуется незамедлительное введение тиамина и, как правило, эффективное лечение. В литературе сообщается о широком диапазоне терапевтических подходов и доз тиамина от 1,5 до 600 мг / день , с 10–20 мг / день в виде разделенных доз в течение нескольких недель от легкой полиневропатии и 20–30 мг / день. от умеренной до тяжелой, обычно до исчезновения симптомов. Как правило, дефицит тиамина купируеся с помощью доз 5–30 мг / сут внутривенно (в / в) или внутримышечно (в / м) три раза в день, затем 5–30 мг / сут перорально до исчезновения симптомов. Однако этот подход заметно менее эффективен для людей с хроническими формами нарушений, связанных с дефицитом тиамина, включая энцефалопатии или дефициты TPK1. В последнем случае стоило бы исследовать лечение непосредственно с помощью TPP; однако неясно, будет ли эта форма фосфорилированного тиамина преодолевать гематоэнцефалический барьер и / или достигать субклеточных мишеней, таких как PDH.
Ряд исследований показал обратную связь между уровнями тиамина и симптомами депрессии у взрослых. Исследование показало, что симптомы депрессии значительно улучшились у пациентов с большой депрессией после 6 недель приема тиамина по сравнению с плацебо. Эффекты от приема тиамина могут быть значительными в качестве паллиативного лечения при послеродовой депрессии и играть важную роль в последующем когнитивном развитии ребенка. PPD ассоциируется с повышенным риском развития неспособности к обучению, синдрома дефицита внимания / гиперактивности (ADHD) и тревожных расстройств у детей младшего возраста, что делает PPD критической проблемой как для матери, так и для младенца. Следовательно, добавки с тиамином могут в некоторой степени улучшить углеводный обмен, функцию митохондрий и выработку энергии в мозге.
Источник