Витамин предшественник гормона кальцитриола
Физиология паратгормона настолько сильно связана с витамином D и метаболизмом костной ткани, что рассматривать эти вопросы по отдельности, не потеряв их сути, невозможно. Графически взаимоотношения представлена на рисунке ниже. Паратгормон в первую очередь регулирует содержание ионов кальция в межклеточном пространстве. Витамин D контролирует всасывание кальция из пищи, а также косвенно отвечает за минерализацию костной ткани, которая содержит в себе 99% всего кальция организма.
«Можно ли считать кость органом?» Да! Поскольку костная ткань проявляется очень высокую метаболическую активность, а также содержит большие запасы кальция, она напрямую включена в процессы регуляции содержания кальция в межклеточном пространстве и плазме крови. Нормальная концентрация кальция в межклеточном пространстве необходима для выполнения множества клеточных функций, среди которых передача сигнала между клетками, секреция гормонов, нормальная работа мышц и нервов, поэтому необходимо строгое ее сохранение в определенных пределах.
Гипокальциемия ведет к нервно-мышечной гипервозбудимости; умеренное снижение уровня кальция проявляется гиперестезиями с положительными симптомами Хвостека и Труссо; тяжелая гипокальциемия ведет к развитию тетании, судорог и смерти. Гиперкальциемия проявляется летаргией, слабостью, комой и, наконец, смертью. Около 50% внеклеточного кальция находится в ионизированной форме, но точное соотношение зависит от pH. Содержание кальция внутри клеток на порядки меньше, чем вне ее, но некоторые органеллы, например, митохондрии, содержат в себе повышенные концентрации кальция. Колебания уровня внеклеточного ионизированного кальция обычно не превышают ± 10%.
а) Физиология обмена паратгормона. Паратгормон (паратиреоидный гормон, ПТГ, РТН) по химическому строению является 84-аминокислотным полипептидом; было выяснено, что за биологическую активность гормона ответственны первые 34 аминокислотных остатка паратгормона, именно они используются в клинической практике, например, в лечении остеопороза. Определение последовательности классического рецептора ПТГ показало его связь с G-белком и наличие семи трансмембранных сегментов, проявляющих одинаковое сродство как с паратгормоном, так и с ПТГ-родственным пептидом (ПТГ-рП).
Но ПТГ-рП не обладает таким же активирующим действием на 1-гидроксилазы почек, как сам ПТГ.
Наибольшее количество рецепторов к ПТГ находится в костной ткани и в почках, хотя в других органах они также присутствуют в меньших количествах. Недавно был обнаружен новый подвид ПТГ-рецептора, который связывается с карбоксильным остатком паратгормона. Ранее считалось, что данные рецепторы находятся в неактивном состоянии. Этот недавно обнаруженный рецептор не связывается с ПТГ-рП. Неизвестно, играет ли он какую-либо роль в метаболизме костной ткани или имеет какие-либо другие функции.
Клетки паращитовидных желез также экспрессируют рецепторы, чувствительные к кальцию. И они являются рецепторами, связанными с G-белками, и имеют в своем составе семь трансмембранных доменов. Данные рецепторы могут соединяться с различными катионами, но физиологически подходящими для них являются только двухвалентные катионы кальция и магния. Снижение уровня ионизированного кальция ведет к повышению секреции ПТГ, повышение уровня ионизированного кальция ведет к снижению секреции ПТГ.
Таким образом, и концентрация ионизированного кальция, и концентрация ПТГ в плазме крови колеблются лишь в узких пределах. Были описаны различные мутации данных рецепторов, некоторые из них приводят к повышению их активности, другие — к понижению. Данные мутации являются ключом к пониманию семейной гипокальциурической гиперкальциемии и некоторых гипокальциемических синдромов, например, семейного гипопаратиреоидизма.
Паратгормон (ПТГ) повышает резорбцию кальция в почках, усиливает резорбцию костной ткани, повышает активность D1-гидроксилазы почек. Эти механизмы помогают восстановить уровень кальция в плазме крови.
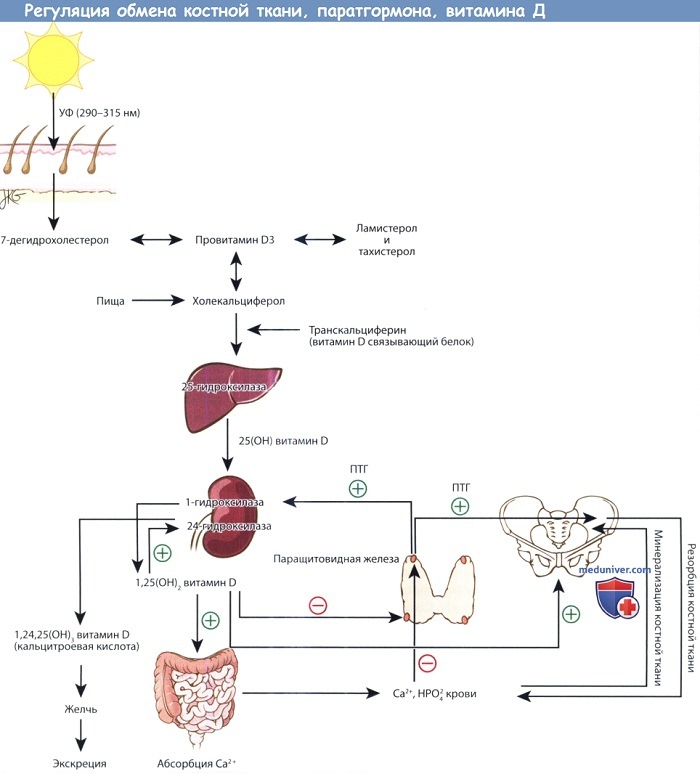 Пути метаболизма витамина D, паратгормона и костной ткани.
Пути метаболизма витамина D, паратгормона и костной ткани.
Предшественники витамина D синтезируются в коже под действием ультрафиолета. Превращение 25-ОН витамина D, депонированной формы витамина, в активную форму,
т.е. 1,25-(ОН)2 витамин D, регулируется паратгормоном. 1,25-(ОН)2 витамин D повышает всасывание кальция в желудочно-кишечном тракте.
Кальций и фосфор плазмы крови регулируют содержание паратгормона крови, а также участвуют в минерализации новообразованного костного матрикса.
При повышении уровня паратгормона усиливается резорбция костной ткани, поскольку это необходимо для поддержания необходимого уровня кальция в плазме,
а также стимулируется синтез 1,25-(ОН)2 витамина D почками.
б) Физиология обмена витамина Д. Синтез провитамина D (холекальциферола) происходит в коже из 7-дегидрохолестерола в результате фотокатализа под действием ультрафиолетовых лучей с длиной волны 290-315 нм. Ультрафиолетовые лучи именно с такой длиной волны могут преодолеть атмосферу, поэтому у жителей возвышенностей синтез провитамина D подвержен сезонным колебаниям. Синтез предшественников провитамина D тоже зависит от воздействия солнечных лучей, поэтому избыточное пребывание на солнце не приводит к гипервитаминозу.
Следовательно, скорость продукции провитамина D в долгосрочной перспективе не зависит от пигментации кожи; но она может снижаться у темнокожих лиц, которые мало подвергаются воздействию солнечных лучей. Провитамин D связывается с транскальциферином (витамин — D — связывающим белком) и транспортируется в печень, где в результате 25-гидроксилирования он превращается в кальцидиол.
Возможности получения витамина D с пищей очень ограничены, лишь крайне малое число продуктов содержит в себе витамин D. Молоко и молочные продукты, производимые в США и Европе, дополнительно обогащаются витамином D. Одна их порция содержит около 100 ME витамина. К сожалению, необогащенные молочные продукты и человеческое грудное молоко содержат крайне малое количество витамина. В желтке яйца содержится около 20 ME витамина D, в белке витамина D нет. Рыба является источником, богатым витамином D.
В порции консервированного тунца содержится 250 ME витамина, а в одной порции свежевыловленного лосося может содержаться до 1000 ME. В растительных продуктах витамина D нет, но он может содержаться в грибах в количестве до 1500 МЕ/100 г, если грибы росли в присутствии солнечного света.
Витамин D представляет из себя провитамин, активной формой которого является 1-25-(ОН)2 витамин D (кальцитриол). В норме кальцитриол продуцируется почками из витамина D, кальцидиола, в результате реакции 1-гидроксилирования. Активность 1-гидроксилазы почек контролируется ПТГ, поэтому уровень 1-25-(ОН)2 витамина D напрямую коррелирует с уровнем ПТГ. Высокий уровень ПТГ, как правило при гипокальциемии, стимулирует образование 1-25-(ОН)2 витамина D. В результате происходит повышение абсорбции кальция в кишечнике и нормализация его уровня в плазме крови.
При определенных патологических состояниях некоторые другие ткани также могут проявлять 1-гидроксилазную активность, иногда независимо от действия ПТГ.

в) Метаболизм минеральных веществ в костной ткани. Мы часто воспринимаем кости лишь как опорные структуры, но на самом деле костная ткань играет важную роль в метаболизме, выступая в качестве депо кальция, фосфатов и карбонатов, также она участвует в поддержании кислотно-щелочного равновесия. Матриксом кости является частично минерализованная ткань и специфические костные клетки. Костную систему человека разделяют на осевой скелет, к которому относят череп, позвоночный столб, грудину, ребра и таз, и на скелет конечностей, от их проксимальных отделов до кончиков пальцев.
Также костная ткань подразделяется на губчатую (трабекулярную), которая обладает высокой метаболической активностью, и кортикальную, которая имеет большую плотность и меньшую метаболическую активность. Изменения в кортикальной костной ткани наступают при тяжелых или длительно текущих заболеваниях. В осевом скелете преобладает содержание трабекулярной кости, в скелете конечностей — кортикальной.
К костным клеткам относят остеобласты, остеокласты и остеоциты. Остеобласты происходит из мезенхимальных стволовых клеток, они образуют соединительнотканный матрикс кости, который затем минерализуется, формируя новую костную ткань. Большинство остеобластов подвергаются апоптозу, но часть из них остается включенной в костный матрикс и превращается в остеоциты, а другая часть остается на поверхности кости, формируя надкостницу.
Трансформируясь в остеоциты, эти клетки образуют длинные отростки, которыми они создают щелевой контакт с соседними клетками и с клетками надкостницы. Остеоциты воспринимают механическое давление, оказываемое на кость, и играют важную роль в моделировании костной ткани. Остеокласты представляют собой крупные многоядерные клетки, происходящие из одноядерных макрофагов. Процесс дифференцировки клеток в остеокласты происходит под контролем ядер-ного фактора кВ (каппа-Б, RANKL), продуцирующего в ходе апоптоза остеобластов, и под контролем колониестимулирующего фактора макрофагов.
Остеокласты участвуют в резорбции костной ткани, разрушая минеральный компонент и коллагеновый матрикс костной ткани посредством протеолиза.
В норме костная ткань постоянно разрушается остеокластами и одновременно заново синтезируется остеобластами. Этот процесс получил название ремоделирования костной ткани. В здоровом организме процессы разрушения и синтеза костной ткани находятся в равновесии. Ремоделирование костной ткани крайне важно для поддержания структурной целостности (прочности) скелета, т. к. с ее помощью постоянно восстанавливаются микропереломы костей, возникающие от каждодневных физических нагрузок.
г) Кальций-чувствительные рецепторы. Как уже упоминалось выше, паращитовидные клетки экспрессируют кальций-чувствительные рецепторы. Первоначально данные рецепторы были выделены из паращитовидных клеток крупного рогатого скота, затем их наличие было подтверждено и у человека. Кроме паращитовидных желез, где они и были впервые обнаружены, данные рецепторы экспрессируются в почках, костях, желудке, легких, головном мозге и других тканях.
Как мутации, так и приобретенные дисфункции данных рецепторов могут стать причиной появления различных заболеваний, связанных как с повышением, так и с понижением кальция крови. Более подробно эти заболевания будут рассмотрены ниже.
Учебное видео расшифровки биохимического анализа крови
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Источник
Витамин предшественник гормона кальцитриола
Кальцитриол является активным метаболитом витамина D3 и в отличие от двух других кальцийрегулирующих гормонов — паратирина и кальцитонина — имеет стероидную природу.
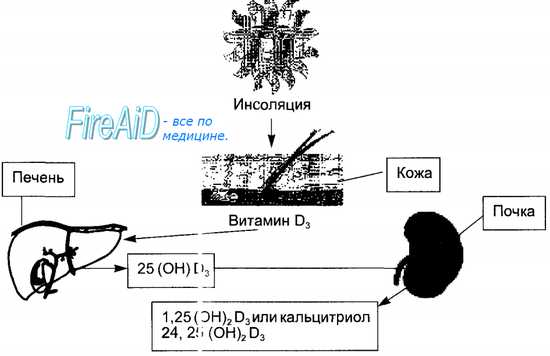 Рис. 6.24. Схема образования к, кальцитриола или активной формы витамина D3. Под воздействием ультрафиолетовых лучей в коже из холестерина образуется витамин D3; поступая с кровью в печень, он подвс и ается первому гидроксилированию по 25-му атому углерода, затем из печени с кровью попадает в почки, где подвергается второму гидроксилированию по 1-му атому углерода, что и ведет к образованию дважды гидроксилированного витамина или кальцитриола.
Рис. 6.24. Схема образования к, кальцитриола или активной формы витамина D3. Под воздействием ультрафиолетовых лучей в коже из холестерина образуется витамин D3; поступая с кровью в печень, он подвс и ается первому гидроксилированию по 25-му атому углерода, затем из печени с кровью попадает в почки, где подвергается второму гидроксилированию по 1-му атому углерода, что и ведет к образованию дважды гидроксилированного витамина или кальцитриола.
Синтез кальцитриола происходит в три этапа (рис. 6.24). Первый этап протекает в коже, где под влиянием ультрафиолетовых лучей из провитамина образуется витамин D3 или холекальциферол. Второй — связан с печенью, куда холека льциферол транспортируется кровью и где в эндоплазматическом ретикул ‘ме гепатоцитов происходит его гидроксилирование по 25-му атому углерода с образованием 25(OH)D3. Этот метаболит поступает в кровь и циркулирует в связи с альфа-глобулином. Его физиологические концентрации не влияют на обмен кальция. Третий этап осуществляется в почках, где в митохондриях клеток проксимальных канальцев происходит второе гидроксилирование и образуются два соединения: l,25-(OH)2-D и 24,25-(OH)2-D. Первое — является наиболее активной формой витамина D3, обладает мощным регуляторным влиянием на обмен кальция в организме и называется кальцитриолом. Образование в почках этого гормона регулируется паратирином, который стимулирует гидроксилирование по первому атому углерода. Таким же эффектом обладает и гипокальциемия. При избытке кальция в крови гидроксилирование происходит по 24-му атому углерода и синтезируется второе соединение — 24,25-(OH)2-D, которое обладает способностью угнетать секрецию паратирина по принципу обратной связи. Инактивация кальцитриола происходит в печени.
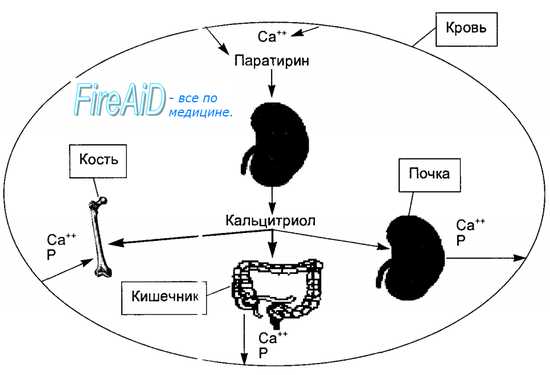 Рис. 6.25. Основные эффекты кальцитриола. Под влиянием паратирина почка секретирует кальцитриол, основные эффекты которого (жирные стрелки) заключаются в стимуляции всасывания в кишечнике в кровь ионов кальция и фосфата и усилении их захвата костной тканью. Эффекты кальцитриола на почку (стимуляция реабсорбции кальция и фосфора) выражены слабее.
Рис. 6.25. Основные эффекты кальцитриола. Под влиянием паратирина почка секретирует кальцитриол, основные эффекты которого (жирные стрелки) заключаются в стимуляции всасывания в кишечнике в кровь ионов кальция и фосфата и усилении их захвата костной тканью. Эффекты кальцитриола на почку (стимуляция реабсорбции кальция и фосфора) выражены слабее.
Основной эффект кальцитриола (рис. 6.25) заключается в активации всасывания кальция в кишечнике. Гормон стимулирует все три этапа всасывания: захват ворсинчатой поверхностью клетки, внутриклеточный транспорт, выброс кальция через базолатеральную мембрану во внеклеточную среду. Действие кальцитриола на эпителиальные клетки кишечника состоит в индуцировании синтеза энтероцитами специальных кальций-связывающих и транспортирующих белков — кальбайндинов. Кальцитриол повышает в кишечнике и всасывание фосфатов. Почечные эффекты гормона заключаются в стимуляции реабсорбции фосфата и кальция канальцевым эпителием. Эффекты кальцитриола на костную ткань связаны с прямой стимуляцией остеобластов и обеспечением костной ткани усиленно всасывающимся в кишечнике кальцием, что активирует рост и минерализацию кости.
Эффекты кальцитриола, как и всех стероидных гормонов, делятся на геномные и негеномные. Геномные эффекты обусловливают синтез кальбайндинов, активирование остеобластов и синтеза костной ткани. Увеличение транспорта кальция внутрь клеток, например в скелетных и сердечной мышцах, в остеобластах, энтероцитах, гепатоцитах и клетках околощитовидных желез, происходит быстро и обусловлено негеномным действием гормона. Под влиянием гормонрецепторного мембранного комплекса в клетках происходит образование вторичного посредника диацил-глицерола и активация протеинкиназы С. Кальцитриол меняет в клетке и уровень цАМФ и цГМФ, что ведет к модификации геномного эффекта. Наличие специфических рецепторов к гормону у многих клеток тканей (в молочной железе, эндокринных железах, нервной системе), способность кальцитриола активировать транспорт кальция в большинстве из них свидетельствуют о широком спектре эффектов этого гормона.
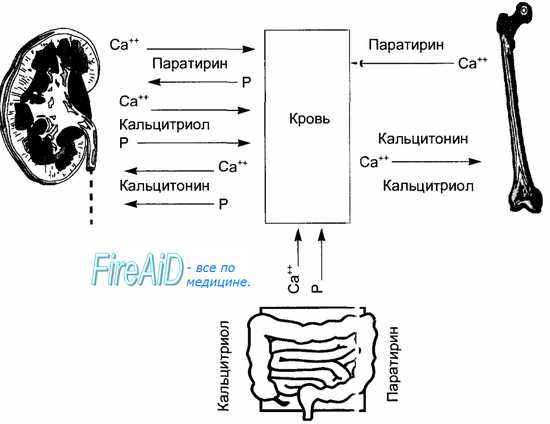 Рис. 6.26. Эффекты трех кальцийрегулирующих гормонов на органы-мишени. В почке паратирин и кальцитриол активируют реабсорбцию кальция, а кальцитонин ее угнетает. И паратирин, и кальцитонин подавляют реабсорбцию фосфата. В кишечнике кальцитриол и паратирин активируют всасывание кальция и фосфата. Кальцитонин и кальцитриол способствуют отложению кальция в костях, а паратирин активирует резорбцию кости и выход кальция в кровь.
Рис. 6.26. Эффекты трех кальцийрегулирующих гормонов на органы-мишени. В почке паратирин и кальцитриол активируют реабсорбцию кальция, а кальцитонин ее угнетает. И паратирин, и кальцитонин подавляют реабсорбцию фосфата. В кишечнике кальцитриол и паратирин активируют всасывание кальция и фосфата. Кальцитонин и кальцитриол способствуют отложению кальция в костях, а паратирин активирует резорбцию кости и выход кальция в кровь.
Участие трех кальций регулирующих гормонов в гомеостазисе кальция и фосфора показано на рис. 6.26.
Недостаточность кальцитриола проявляется в виде рахита, т. е. нарушения созревания и кальцификации хрящей и кости у детей, либо остеомаляции, т. е. падения минерализации костей после завершения роста скелета. При этом сдвиги уровня кальция в крови и клетках обусловливают угнетение нейромышечной возбудимости и мышечную слабость.
Источник