Параграф 46 синтез жирных кислот
Автор текстов Анисимова Елена Сергеевна.
Авторские права защищены. Продавать текст нельзя.
Курсив не зубрить.
Замечания присылать по электронной почте exam_bch@mail.ru
https://vk.com/bch_5
ПАРАГРАФ 46:
«СИНТЕЗ ЖИРНЫХ КИСЛОТ».
См. формулы в файле «46 формулы».
46.1. ЗНАЧЕНИЕ синтеза жирных кислот для организма –
этот процесс является источником жирных кислот для нужд клеток.
Основной путь использования жирных кислот в организме –
использование жирных кислот для синтеза жира (в основном в качестве «резерва энергии» на случай голода),
а также для синтеза липоидов, в том числе для синтеза МЕМБРАННЫХ липидов,
что нужно для образования новых клеток (для деления) и для «ремонта» мембран – то есть для замены молекул мембран новыми неповреждёнными молекулами (иначе мембраны быстрее разрушались бы, что приводило бы к гибели клеток).
Подробнее о том, зачем нужны жир и липоиды – см. п.47 и …
Снижение синтеза жирных кислот может привести к дефициту в организме липоидов, что в свою очередь может привести к дерматитам и другим проявлениям снижения пролиферативных процессов (деления клеток).
При снижении синтеза жирных кислот остаётся такой источник жирных кислот, как поступление жирных кислот с пищей в составе жира (точнее, в виде ацилов в составе пищевых липидов).
ГДЕ синтезируется пальмитиновая кислота.
Основное место синтеза пальмитиновой кислоты – печень. Не жировая ткань, не адипоциты, а печень.
После синтеза в печени пальмитиновая кислота используется для синтеза молекул жира (см. …) и липоидов, которые включаются в состав липопротеинов очень низкой плотности (ЛПОНП), которые поступают в кровь для доставки липидов в ткани. Подробности обмена ЛПОНП смотрите в п. … В итоге синтезированные в печени жирные кислоты оказываются в жировой ткани в составе жира – в качестве резерва энергии на случай голода.
46.2. Какие жирные кислоты синтезируются в организме человека.
В основном в организме синтезируется пальмитиновая кислота (её соли и анионы называются пальмитатами) с 16-тью атомами углерода и формулой С16Н31СООН (обозначение – С16:0).
Из неё может синтезироваться стеариновая кислота (С18:0), а из стеариновой – олеиновая (С18:1) и эйкозановая (С20:0).
Из линолевой кислоты может синтезироваться арахидоновая кислота (АрК).
Из линоленовой кислоты может синтезироваться эйкозапентаеновая (ЭПЕ) а из неё – докозагексаеновая (ДГЕ).
При этих реакциях происходит удлинение углеродной цепочки жирных кислот, которое называется элонгацией.
При образовании олеиновой кислоты из стеариновой, а также АрК из линолевой и ЭПЕ из линоленовой, ДГЕ из ЭПЕ происходит образование новых двойных связей, которое называется десатурацией и катализируется ферментами десатуразами.
Линолевая и линоленовая кислоты не могут синтезироваться в организме, должны поступать с пищей и поэтому называются незаменимыми жирными кислотами – аналогично тому как есть незаменимые аминокислоты.
Незаменимые жирные кислоты поступают в организм благодаря присутствию в пище растительных масел.
АрК, ЭПЕ и ДГЕ кислоты не считаются незаменимыми жирными кислотами, но лучше, если они поступают в организм с пищей (морепродукты).
При отсутствии в пище растительных масел и морепродуктов организм не получает незаменимы жирных кислот, что приводит к снижению синтеза мембранных липидов и деления клеток.
46.3. Регуляция синтеза жирных кислот.
Регуляция гормонами. Синтез жирных кислот происходит в состоянии сытости и покоя под влиянием инсулина (количество которого в крови увеличивается в этом состоянии).
В состоянии голода синтез жирных кислот снижается (по поговорке «не до жиру – быть бы живу») под влиянием гормона голода глюкагона.
В состоянии стресса и при работе синтез жирных кислот также снижается – под влиянием гормонов стресса катехоламинов (адреналина и норадреналина).
Дополнительно. – Синтез жирных кислот усиливается женскими половыми гормонами эстрогенами.
Регуляция метаболитами. Как обычно, продукты процесса и их метаболиты замедляют процесс, а субстраты усиливают (ацетилКоА).
46.4. Реакции синтеза пальмитиновой кислоты.
Синтез пальмитиновой кислоты осуществляется комплексом ферментов, который называется синтазой жирных кислот.
Из чего синтезируется молекула пальмитиновой кислоты – из молекул ацетилКоА. Источник ацетилКоА для синтеза жирных кислот – образование ацетилКоА из глюкозы (в ходе гликолиза и ПДГ – см. …). Именно поэтому сладости, мучное и картофель способствуют набору веса.
Для синтеза одной молекулы пальмитиновой кислоты (16 атомов углерода) нужно 8 молекул ацетилКоА (одна молекула ацетилКоА – это 2 атома углерода), атомы углерода которых нужно соединить в «цепочку».
Присоединение атома углерода каждого очередного ацетила происходит по очереди. К первому ацетилу присоединяется второй ацетил и т.д., пока не соединятся все 8 молекул, образовав молекулы пальмитиновой кислоты.
Далее идёт описание реакций. Формулы веществ и реакции смотрите в таблице к 46-му параграфу. Здесь только словесное описание формул и реакций таблицы. Без иллюстрации в таблице не поймёте.
1. «МалонилКоА». Семь молекул ацетилКоА из восьми превращаются в малонилКоА за счёт реакции с СО2 (точнее, с бикарбонатом – НСО3 – , который образуется из СО2), при которой образуется карбоксильная группа (из-за чего реакция называется карбоксилированием ацетилаКоА, а фермент, катализирующий эту реакцию, называется карбоксилазой ацетила КоА или ацетилКоА/карбоксилазой).
Для этой реакции необходим витамин Н (БИОТИН), поэтому при отсутствии биотина синтез жирных кислот не происходит, а дефицит биотина проявляется дерматитами (причиной дефицита биотина может быть дисбактериоз).
СО2 присоединяется ко второму атому углерода ацетилКоА.
Реакция происходит с затратой АТФ – АТФ расщепляется на АДФ и фосфат для выделения энергии.
По одной АТФ на синтез каждой молекулы малонилКоА, то есть всего тратится 7 молекул АТФ.
Сравните реакцию с карбоксилированием пирувата – много похожего.
Синтез жирных кислот – пример анаболического процесса, требующего затрат энергии.
2. «Замена КоА на АПБ, перенос с КоА на АПБ».
Первая молекула ацетилКоА и 7 молекул малонилКоА вступают в реакцию с АПБ (ацилпереносящим белком) для того, чтобы отсоединиться от КоА и присоединиться к АПБ. В результате образуются: 1 молекула ацетилАПБ и 7 молекул малонилАПБ.
3. Первая молекула ацетилАПБ вступает в реакцию с первой (с одной из семи) молекул малонилАПБ.
При этом соединяются первый атом углерода ацетилАПБ и ВТОРОЙ атом углерода малонил АПБ,
образуя молекулу из 4 атомов углерода с кетогруппой в третьем (;) положении, которая называется ;-кето/бутирилАПБ.
В реакции первый атом углерода ацетилАПБ отсоединяется от АПБ, а –СООН (карбоксильная группа) отсоединяется от второго атома углерода малонилаКоА и выделяется в реакции в виде СО2. (Таким образом, СО2 на синтез не тратится).
Бутирил – это остаток (ацил) 4-хуглеродной жирной кислоты (бутановой).
Далее все остатки любой длины называются просто ацилами.
У ;-кетобутирилАПБ есть кетогруппа. Нужно сделать так, чтобы на её месте в ;-положении была –СН2– группа (метиленовая), то есть нужно восстановить кетогруппу по метиленовой. Это происходит благодаря трём реакциям, «противоположным» реакциям ;-окисления.
4.1. Кетогруппа ;-кетобутирилаАПБ превращается в гидроксильную, в результате чего ;-КЕТОбутирилАПБ превращается в ;-ГИДРОКСИбутирилАПБ.
При этом происходит присоединение двух атомов водорода, источником которых является, как обычно, НАДФН, Н+, для образования которого клеткам нужен витамин РР. Катализируют реакции такого типа (перенос водорода от НАДФН на субстрат) РЕДУКТАЗЫ. Редуктаза данной реакции называется редуктазой ;-кетобутирилаАПБ или ;-кетобутирилАПБ/редуктазой.
Источником НАДФН, Н+ являются реакции ПЕНТОЗОФОСФАТНОГО пути (ПФП; вариант «пентозный цикл» в данном случае) – см. … , для протекания которого нужны ГЛЮКОЗА в качестве субстрата и ИНСУЛИН в качестве гормона, стимулирующего ПФП, а также витамины РР и В1.
4.2. ;-ГИДРОКСИбутирилАПБ теряет молекулу воды (НОН), при этом ОН отщепляется от 3-го атома углерода, а атом водорода (Н) – от второго. В итоге образуется вещество с двойной связью между 2-м и 3-и атомами углерода (; и ;), которое называется ЕНоилАПБ. (ЕН означает двойную связь).
Процесс отщепления Н и ОН и виде воды называется дегидратаций (не путать с дегидрированием), а фермент, который катализирует дегидратацию, называется дегидратазой, дегидратаза гидрокси/бутирилаАПБ называется гидрокси/бутирилАПБ/дегидратазой.
4.3. К еноилуАПБ по двойной связи присоединяются 2 атома водорода, в результате чего образуется бутирилАПБ.
Источником 2 атомов водорода является, как обычно, НАДФН, Н+. Катализируют реакции такого типа (перенос водорода от НАДФН на субстрат) РЕДУКТАЗЫ. Редуктаза данной реакции называется редуктазой еноилаАПБ или еноилАПБ/редуктазой.
БутирилАПБ является ацилом с 4 атомами углерода. Далее он вступает в реакцию с очередной (второй из семи исходных и из шести оставшихся) молекулой малонилАПБ так же, как вступал ацетилАПБ. – То есть первый атом углерода бутирилАПБ соединяется со вторым атомом углерода малонилаАПБ, АПБ и СО2 «уходят», образуется соединение с 6 атомами углерода и кетогруппой в ;-положении. Далее происходят реакции, аналогичные реакциям 4.1-4.3, в результате которых образуется ацилАПБ с шестью атомами углерода. То есть углеродная цепочка бутирила удлиняется ещё на 2 атома углерода.
АцилАПБ с шестью атомами углерода вступает в реакцию с очередной (то есть третьей из исходных семи) молекул малонилАПБ и т.д.
И так до тех пор, пока не будут использованы все 7 молекул малонилАПБ.
В результате образуется ацил из 16 атомов углерода – пальмитоилАПБ.
После этого остаётся отщепить пальмитоил от АПБ и превратить его в пальмитиновую кислоту.
5. ПальмитоилАПБ превращается в пальмитиновую кислоту за счёт расщепления связи между пальмитоилом и АПБ путём гидролиза (то есть с присоединением воды «по местам расщепления связи»).
46.4
Синтез жирных кислот – пример типичного анаболического процесса. (То есть процесса синтеза).
При этих процессах из простых веществ образуются более сложные – в данном случае 16-тиуглеродная цепочка из двухуглеродных молекул ацетилКоА.
Анаболические процессы протекают с затратой АТФ (7 молекул на одну молекулу пальмитиновой кислоты) и НАДФН (7*2= 14 молекул на одну молекулу пальмитиновой), поставляемого пентозофосфатным путём.
46.5
Сравнение синтеза жирных кислот и ;-окисления –
1. Синтез – процесс анаболический, а ;-окисление – катаболический.
2. АТФ: при синтезе АТФ тратится (7), а при ;-окислении образуется (108 АТФ за счёт одного пальмитата).
3. Коферменты: при синтезе НАДФН (образующийся в ПФП) превращается в НАДФ+ (который снова превращается в НАДФН в ПФП),
а при ;-окислении – НАД+ и ФАД и превращаются в НАДН и ФАДН2, поступающие в дыхательную цепь для выработки энергии.
4. Витамины: для синтеза нужны биотин и РР, а для ;-окисления – РР и В2. В составе коферментов.
5. Переносчики ацилов: при синтезе кофермент А (КоА) и АПБ (ацилпереносящий белок), а при ;-окислении – только кофермент А.
6. Локализация – синтез в печени, а ;-окисление – во всех клетках, кроме эритроцитов и нейронов.
7. В каких условиях протекают: синтез при сытости и покое под влиянием инсулина,
а ;-окисление – при стрессе под влиянием катехоламинов адреналина и норадреналина и при голоде под влиянием гормона голода глюкагона.
8. Какими гормонами стимулируются: синтез – инсулином и эстрогенами, а ;-окисление – катехоламинами и глюкагоном.
9. Какими гормонами тормозятся: синтез – катехоламинами при стрессе и глюкагоном при голоде, а ;-окисление – глюкагоном при голоде.
Источник
Синтез жирных кислот
Субстраты синтеза жирных кислот. Предшественником является ацетил-КоА, процесс протекает в цитозоле (рис. 13.3). Ацетил-КоА образуется из пирувата под действием митохондриального пируватдегидрогеназного комплекса. Внутренняя митохондриальная мембрана непроницаема для ацетил-КоА. В митохондриях фермент цитратсинтаза катализирует реакцию образования цитрата из ацетил-КоА и ЩУК. Цитрат выходит из митохондрий в цитоплазму. В цитозоле фермент АТФ-цитратлиаза расщепляет цитрат до ацетил-КоА и ЩУК.
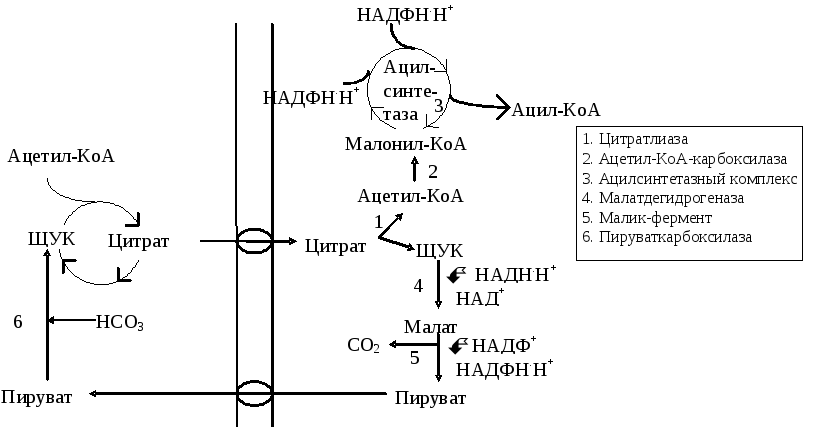
Рис. 13.3. Субстраты для синтеза жирных кислот. Переход ацетил-КоА из митохондрий в цитозоль
Ферменты синтеза жирных кислот. В биосинтезе насыщенных жирных кислот участвуют два ферментных комплекса: ацетил-КоА карбоксилаза и ацилсинтетаза (рис. 13.4).

Рис. 13.4. Схема реакций синтеза жирных кислот
Ацилсинтетазный мультиферментный комплекс. Комплекс содержит ацилпереносящий белок (АПБ) в качестве своеобразного ядра. Другими ферментами комплекса являются -кетоацилсинтетаза (КС), малонилтрансфераза (МТ), -кетоацилредуктаза (КР), -гидро-ксиацилдегидратаза (ГД), еноилредуктаза (ЕР) и ацилтрансацетилаза (АТ).
Ацетил-КоА карбоксилаза катализирует ключевую реакцию в синтезе жирных кислот:

Цитрат активирует фермент. Ацетил-КоА карбоксилаза подвергается обратимому фосфорилированию/дефосфорилированию; цАМФ-зависимая протеинкиназа ингибирует ферментативную активность, а фосфатаза — активирует. На этом основана гормональная регуляция активности ацетил-КоА карбоксилазы. Повышенные концентрации малонил-КоА и ацил-КоА аллостерически ингибируют активность этого фермента.
За 7 таких циклов образуется насыщенная жирная кислота с 16 углеродными атомами. В общем виде это выглядит следующим образом:
Потребность в НАДФН . Н + для реакций восстановления в процессе синтеза жирных кислот. Источником восстановленного НАДФ + является окислительный этап пентозофосфатного пути обмена глюкозы, а также реакции переноса водорода в пути расщепления цитрата. Последние протекают с участием фермента малатдегидрогеназы (кофермент НАДН . Н + ) и малик-фермента (в реакции восстановления НАДФ + ).
ПРОИСХОЖДЕНИЕ НЕНАСЫЩЕННЫХ ЖИРНЫХ КИСЛОТ В КЛЕТКАХ
Незаменимые и заменимые. В организме человека не могут синтезироваться -3 и -6 жирные кислоты в связи с отсутствием ферментной системы, которая могла бы катализировать образование двойной связи в положении -6 или любом другом положении, близко расположенном к -концу. К таким жирным кислотам относятся: линолевая кислота (18:2, 9, 12), линоленовая кислота (18:3, 9, 12, 15), арахидоновая кислота (20:4, 5, 8, 11, 14). Арахидоновая кислота является незаменимой только при недостатке линолевой кислоты.
Функции полиненасыщенных жирных кислот: придают жидкостность мембранам, являются предшественниками эйкозаноидов
Э йкозаноиды (липидные гормоны). Простагландины, тромбоксаны и лейкотриены. Самым главным предшественником является арахидоновая кислота.
йкозаноиды (липидные гормоны). Простагландины, тромбоксаны и лейкотриены. Самым главным предшественником является арахидоновая кислота.
Схема высвобождения арахидоновой кислоты из мембранных фосфолипидов:
 Судьба арахидоновой кислоты, которая высвободилась из состава мембранных фосфолипидов:
Судьба арахидоновой кислоты, которая высвободилась из состава мембранных фосфолипидов:
Ингибиторы синтеза эйкозаноидов. Кортикостероиды ингибируют фосфолипазу А2, тем самым они ингибируют высвобождение арахидоновой кислоты из состава мембранных фосфолипидов. Ингибиторы из числа противовоспалительных препаратов нестероидной природы (аспирин, индометацин, фенилбутазон) ингибируют циклоксигеназу.
Простагландины стимулируют сокращение гладких мышц, липолиз, снижают секрецию желудочного сока, свертывание крови, вызывают расширение кровеносных сосудов.
Тромбоксаны стимулируют агрегацию тромбоцитов, сокращение стенки артерий, образование цАМФ в тромбоцитах, повышают кровяное давление.
Лейкотриены участвуют в формировании воспаления, аллергических реакций, в хемотаксисе.
Источник