Синтез гема витамин в12
Гемоглобин — это гемопротеин, с молекулярной массой около 60 тыс., окрашивающий эритроцит в красный цвет после связывания молекулы O2 с ионом железа (Fe++). У мужчин в 1 л крови содержится 157 (140—175) г гемоглобина, у женщин — 138 (123—153) г. Молекула гемоглобина состоит из четырех субъединиц гема, связанных с белковой частью молекулы — глобином, сформированной из полипептидных цепей.
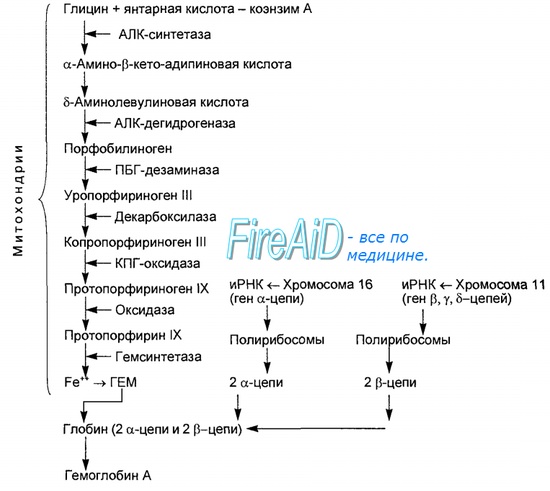
Синтез гема протекает в митохондриях эритробластов. Синтез цепей глобина осуществляется на полирибосомах и контролируется генами 11-й и 16-й хромосом. Схема синтеза гемоглобина у человека представлена на рис. 7.2.
Гемоглобин, содержащий две а- и две В-цепи, называется А-тип (от adult — взрослый). 1 г гемоглобина А-типа связывает 1,34 мл O2. В первые три месяца жизни плода человека в крови содержатся эмбриональные гемоглобины типа Gower I (4 эпсилон цепи) и Gower II (2а и 25 цепи). Затем формируется гемоглобин F (от faetus — плод). Его глобин представлен двумя цепями а и двумя В. Гемоглобин F обладает на 20—30 % большим сродством к O2, чем гемоглобин А, что способствует лучшему снабжению плода кислородом. При рождении ребенка до 50—80 % гемоглобина у него представлены гемоглобином F и 15—40 % — типом А, а к 3 годам уровень гемоглобина F снижается до 2 %.
Соединение гемоглобина с молекулой 02 называется оксигемоглобином. Сродство гемоглобина к кислороду и диссоциация оксигемоглобина (отсоединения молекул кислорода от оксигемоглобина) зависят от напряжения кислорода (Р02), углекислого газа (РС02) в крови, рН крови, ее температуры и концентрации 2,3-ДФГ в эритроцитах. Так, сродство повышают увеличение Р02 или снижение РС02 в крови, нарушение образования 2,3-ДФГ в эритроцитах. Напротив, повышение концентрации 2,3-ДФГ, снижение Р02 крови, сдвиг рН в кислую сторону, повышение РС02 и температуры крови — уменьшают сродство гемоглобина к кислороду, тем самым облегчая ее отдачу тканям. 2,3-ДФГ связывается с р-цепями гемоглобина, облегчая отсоединение 02 от молекулы гемоглобина.
Увеличение концентрации 2,3-ДФГ наблюдается у людей, тренированных к длительной физической работе, адаптированных к длительному пребыванию в горах. Оксигемоглобин, отдавший кислород, называется восстановленным, или дезоксигемоглобином. В состоянии физиологического покоя у человека гемоглобин в артериальной крови на 97 % насыщен кислородом, в венозной — на 70 %. Чем выраженней потребление кислорода тканями, тем ниже насыщение венозной крови кислородом. Например, при интенсивной физической работе потребление кислорода мышечной тканью увеличивается в несколько десятков раз и насыщение кислородом оттекающей от мышц венозной крови снижается до 15 %. Содержание гемоглобина в отдельном эритроците составляет 27,5—33,2 пикограмма. Снижение этой величины свидетельствует о гипохромном (т. е. пониженном), увеличение — о гиперхромном (т. е. повышенном) содержании гемоглобина в эритроцитах. Этот показатель имеет диагностическое значение. Например, гиперхромия эритроцитов характерна для В|2-дефицитной анемии, гипохромия — для железодефицитной анемии.
Источник
Синтез гема витамин в12
Поражение крови, обусловленное первичным нарушением синтеза гема, вызывают свинец и его соединения. Механизм токсического действия свинца на кровь характеризуется вмешательством его в порфириновый обмен, в частности в процесс синтеза гема, представляющего собой железопорфирин. Биосинтез гема осуществляется в основном в митохондриях эритробластов костного мозга и представляет собой сложный многоступенчатый ферментативный процесс. Как известно, исходным продуктом синтеза гема является глицин вместе с сукцинил КоА. Последний образуется из янтарной кислоты и коэнзима А. При взаимодействии глицина с сукцинил КоА вначале образуется а-амино-кетоадипиновая кислота.
Фактически первый этап синтеза гема — это синтез дельтааминолевулиновой кислоты (АЛК) в янтарно-глициновом цикле при участии фермента синтетазы АЛК при условии, что коферментом этой реакции служит производное витамина В6-пиридоксальфосфат. Следующая ступень синтеза — превращение АЛК в порфобилиноген под влиянием дегидратазы АЛК. Далее через промежуточные продукты синтеза уро- и копропорфирин, после воздействия на последний фермента декарбоксилазы (копрогеназы) образуется непосредственный предшественник гема протопорфирин. Превращение протопорфирина в гем осуществляется благодаря включению в его молекулу двухвалентного железа, причем катализатором этого процесса является фермент гемосинтетаза.
Угнетающее действие свинца на биосинтез гема обусловлено его ингибирующим влиянием на ферменты, катализирующие синтез гема. Сущность такого ингибирующего влияния заключается в том, что свинец блокирует функционально активные центры ферментов: сульфгидрильные (SH), карбоксильные и аминные группы. Особенно характерной является блокада SH-rpyrm ферментов, участвующих в синтезе гема, что, впрочем, свойственно всем «тиоловым» ядам, к которым принадлежит и свинец.
В результате падения активности фермента дегидратазы АЛК образуется ток АЛК, которая в повышенных количествах выделяется с мочой. Одновременно с тормозящим действием свинца на дегидратазу АЛК аналологичное воздействие оказывается и на ферменты декарбоксилазу и гемсинтетазу. Вследствие этого в эритроцитах накапливается избыток уро- и протопофирина. Причем по мере усугубления свинцовой интоксикации соотношение между количеством избыточного количества копро- и протопорфирина смещается в сторону последнего. Такое изменение первоначальной пропорции между копро- и протопорфирином, по-видимому, обусловлено двояким влиянием свинца на порфириногенез.
С одной стороны, уровень протопорфирина возрастает за счет стимулирования на первом этапе декарбоксилирования копропорфирина (фермент копрогеназа) с превращением его в протопорфирин, а с другой стороны, уровень последнего возрастает благодаря торможению процесса включения железа в протопорфириновое кольцо. По мере развития интоксикации стимулирующее влияние на декарбоксилирование копропорфирина снижается, однако дисбаланс в содержании копро- и протопорфирина в крови с преобладанием последнего сохраняется. Это объясняется усиленным выведением копропорфиринов с мочой, тогда как для протопорфирина почечный барьер непреодолим. Следовательно, в целом сдвиг соотношения копропорфирин/протопорфирин объясняется опять-таки двумя механизмами: гиперпродукцией протопорфирина с нарушением его утилизации, а также усиленным выведением копропорфирина с мочой. Помимо традиционного пути образования протопорфирина из копропорфирина, возможно его образование непосредственно из АЛК.
Особенности вмешательства свинца в обмен порфиринов определяет появление ранних признаков его воздействия на организм. Так, самыми ранними диагностическими показателями влияния свинца на систему крови служит появление в моче дельтааминолевулиновои кислоты в сочетании с копропорфиринурией, а также протопорфиринемия. Угнетение биосинтеза гема как такового приводит к дефициту гемоглобина. Поскольку при этом нарушается включение железа в молекулу протопорфирина, в эритробластах и эритроцитах появляется избыток неутилизированного железа (в виде гранул), вследствие чего они превращаются в сидеробласты и сидероциты.
Нарушение синтеза гемоглобина приводит к стимуляции красного ростка костного мозга, что сопровождается ретикулоцитозом и появлением базофильно-зернистых эритроцитов, что свидетельствует об омоложении красной крови. При этом ретикулоцитоз и базофильная пунктация эритроцитов обычно регистрируются параллельно. Существует точка зрения, согласно которой базофильная зернистость имеет протоплазматическую природу и появление ее обусловлено усилением регенераторной активности в сфере эритропоэза. По мнению И.А.Кассирского (1970), базофильная зернистость в эритробластах и эритроцитах является следствием дезинтеграции кислого коллоида, т.е. рибонуклеиновой кислоты, содержащейся в ретикулуме, что служит достоверным доказательством ее костномозгового происхождения.
Примечательно, что увеличение числа базофильно-зернистых эритроцитов не всегда сопровождается анемией так же, как и ретикулоцитоз. Поэтому оба эти симптома являются наиболее ранними признаками изменений в системе красной крови. При этом базофильная зернистость не является строго специфичной для токсического воздействия свинца, появляясь и под влиянием других ядов крови (бензол, мышьяковистый водород, оксид углерода, анилиз), а также нейротоксических веществ (сероводород, сероуглерод, бензин и т.д.), однако наиболее закономерно ее появление при поступлении в организм свинца. Все же чрезвычайная лабильность этого признака — быстрое появление и столь же быстрое исчезновение базофильно-зернистых эритроцитов даже при непрерывном контакте со свинцом отсутствие корреляции с признаками анемии снижает диагностическую ценность базофильной зернистости, если она регистрируется изолированно в отсутствие других показателей свинцовой интоксикации.
Источник
Синтез гема витамин в12

Цианокобаламин (Витамин В12)
CAS номер: 68-19-9
Брутто формула: C63H88CoN14O14P
Внешний вид: порошок темно-красного цвета без запаха
Химическое название и синонимы: Cyanocobalamin, Cyano-5,6-dimethylbenzimidazole-cobalamin; Vitamin B12.
Физико-химические свойства:
Молекулярная масса 1355.38 г/моль
Растворимость: в воде растворим.
pH: aq solns являются нейтральными
Давление паров: незначительное.
Плотность паров: нет.
Скорость испарения: незначительная.
Вязкость: отсутствует.
Температура замерзания / плавления:> 300 ° C
Температура разложения: недоступна.
Растворимость: умеренно растворим в воде.
Химическая стабильность: Стабилен при нормальных температурах и давлениях. Может разлагаться, когда подвергается воздействию света. Условия, которых следует избегать: свет, пыль, избыток тепла.
Несовместимость с другими материалами: сильные окислители, сильные кислоты.
Опасные продукты разложения: окись углерода, оксиды азота, оксиды фосфора, диоксид углерода, оксиды кобальта.
Витамин В12 (Кобаламин) – встречается большей степенью в печени, красном мясе, рыбе и почках. Организм человека может также сам синтезировать Витамин В12 в толстом отделе кишечника с помощью микроорганизмов, но так как всасывается витамин в подвздошной кишке, то польза от аутогенного кобаламина сомнительная.
B12 представляет собой тетрапиррольный комплекс, который содержит кобальт в молекуле и может относиться к нескольким формам кобаламина: цианокобаламин (CN-Cbl) и гидроксокобаламин (OH-Cbl) – эти формы витамина B12 доступны для медицинского использования. Аденозилкобаламин (Ado– Cbl), метилкобаламин Me-Cbl) и кобинамид (CN2-Cbn) такие формы витамина B12 обнаруживаются в биологических или пищевых образцах.
Впервые витамин был открыт в США и в Англии в 1948 году. Позже почти через 30 лет пробовали синтезировать предшественник витамина В12, но из- за сложности данного многостадийного процесса не начал производиться в крупных масштабах. Затем витамин пробовали получать непосредственно из печени, но выход витамина был очень не большой. В настоящее время получают витамин исключительно с помощью микробиологического синтеза.
Роль витамина В12 для организма заключается во множестве функций, включающих участие витамина в синтезе некоторых незаменимых аминокислот, нуклеиновых кислот, миелина (вещество, образующее нервную оболочку), холина, метионина, а также кобаламин стимулирует функцию костного мозга (который отвечает за гемопоэз), способствует образованию гема в эритроцитах. Витамин B12 может участвовать в поддержании сульфгидрильных (SH) групп в восстановленной форме, необходимой для многих SH-активируемых ферментных систем. Благодаря этим реакциям витамин B12 связан с жировым и углеводным обменом и синтезом белка. Дефицит витамина B12 приводит к некоторым видам анемии (например, мегабластной), поражениям пищеварительного тракта и неврологическим патологиям (которые начинаются с неспособности продуцировать миелин и сопровождаются постепенной дегенерацией аксона и нервной головки). Также д ефицит витамина B12 потенциально может привести к серьезным и необратимым повреждениям, особенно мозга и нервной системы. При даже небольшом дефиците В12 , могут наблюдаться такие симптомы, как усталость, вялость, депрессия, плохая память, одышка, головные боли и бледность кожи, в частности, особенно у пожилых людей (старше 60 лет). Дефицит витамина B12 также может вызывать симптомы мании и психоза.
Витамин в12 применяется в медицине и ветеринарии при гипо- и авитаминозах витамина. Существует огромное множество форм витамина в виде таблеток, капсул, пероральных жидкостей, спреев, порошков, в качестве БАДов, в составе мультивитаминных комплексов, в форме растворов для инфузий, даже в составе зубных паст и продуктах питания с целью обогащения пищи витамином. Назначают витамин в12 при различных анемиях, рассеянном склерозе, при недостаточности витаминов группы В, болезнях тройничного нерва и двигательного нейрона, параличе и парезе, при печеночной недостаточности, невралгии и многих других. Часто прием добавок с витамином В12 показан для приверженцев вегетерианского и веганского образа жизни с целью профилактики гиповитаминоза В12 или для устранения авитаминоза В12 и их последствий для организма ,по причине недостаточного потребления продуктов содержащих В12 или неупотребления их вовсе.
Получение витамина в12 осуществляют путем микробиологического синтеза из таких организмов, как актиномиценты, водоросли одноклеточные, а также фото- и метано синтезирующие бактерии. Например, источником В12 служат пропионовокислые бактерии, которых культивируют на средах с глюкозой, неорганическими солями, хлоридом кобальта, казеиновым гидролизатом и витаминами. Затем в среду добавляют 5,6-диметилбензимидазол( предшественник витамина), наблюдается активный рост в течение 5-6 суток и синтез витамина с выходом около 5,6-8,7 мг/л. Для более успешного синтеза и высокого выхода витамина активно также используют кукурузный субстрат и мутант пропионовокислых бактерий, выход витамина может составлять в таком случае до 30 мг/л витамина В12.
Действие на организм:
При пероральном попадании в организм В12 связывается с белком слюны — R-протеином, который защищает его в желудке от действия соляной кислоты. При поступлении в желудок, витамин, освободившись от R-протеина, соединяется с гликопротеидом – внутренним фактором Касла (Кастла) (который в свою очередь, вырабатывается париетальными клетками фундальных желез, расположенных в области дна тела и интрамедиальной части желудка) – проводящего его через агрессивную среду желудочного сока, защищая его от пищеварительных ферментов, а также от микрофлоры кишечника. В соединении с гликопротеидом В12 становится способен проникнуть через эпителиальные клетки подвздошной кишки и далее с помощью транспортных белков крови (альфа- и бетаглобулинов) в виде гидроксикобаламина разносится по организму и главным образом попадает в печень.
Сам по себе кобаламин не активен, для включения себя в два основных процесса — окисления остатка жирных кислот и трансаминирования аминокислоты гомоцистеина с последующим образованием метионина — принимает две коферментные формы: дезоксиаденозин-кобаламин и метилкобаламин соответсвенно.
Дезоксиаденозин-кобаламин участвует в реакции изомеризации – переноса радикальной группы на место водорода, а водорода на свободную связь углерода. В реакциях окисления незаменимых аминокислот (валин, изолейцин, цистеин и т.д.), это приводит к образованию большого количества энергии в виде сукценил-SKoA, используемого в цикле трикарбоновых кислот.
Метилкобаламин помогает образованию метионина из гистидина. Метионин является предшественником таких незаменимых веществ, как – адреналин, креатин, карнитина и др. Кроме того, реакция образования метионина способствует удержанию фолиевой кислоты (Витамин В9) внутри клетки.
Гиповитаминозы проявляются чаще всего не в следствии неправильного питания, а из-за нарушения в одном из звеньев проникновения кобаламина в организм, будь то дефицит внутреннего фактора Касла, поражение эпителиальных клеток нисходящей части подвздошной кишки и прочие нарушения мешающие нормальному транспорту витамина. Как правило, дефицит можно восполнить добавлением в рацион витамина В12 или применением инъекционных форм цианокобаламина (Естественно, это не будет являться лечением основного заболевания в большинстве случаев).
Цианокобаламин вводят интраназально, перорально и парентерально, тогда как гидроксокобаламин вводят только парентерально. Хранится витамин в основном в печени, но в костном мозге также содержится значительное количество поглощенного витамина В12. Этот витамин проникает через плаценту и распространяется в грудное молоко. Энтерогепатическая рециркуляция сохраняет системные запасы. Период полувыведения составляет около 6 суток (а в печени в среднем 400 дней). Ликвидация происходит главным образом через желчь. Однако избыток цианокобаламина из организма экскретируется и в неизмененном виде с мочой.
Из культуральной жидкости выделяют витамин, с помощью экстракции применяя органические растворители, ионообменной хроматографией. Если необходим витамин В12 для животноводческих целей, то продуцентами могут служить метанобразующие бактерии, а для медицинского использования витамин В12 получают обычно из бактерий Propionibacterium shermanii.
LD50 при внутривенном введении — мышь — 2 г / кг
Источник