Синтез эритроцитов дефицит витамина
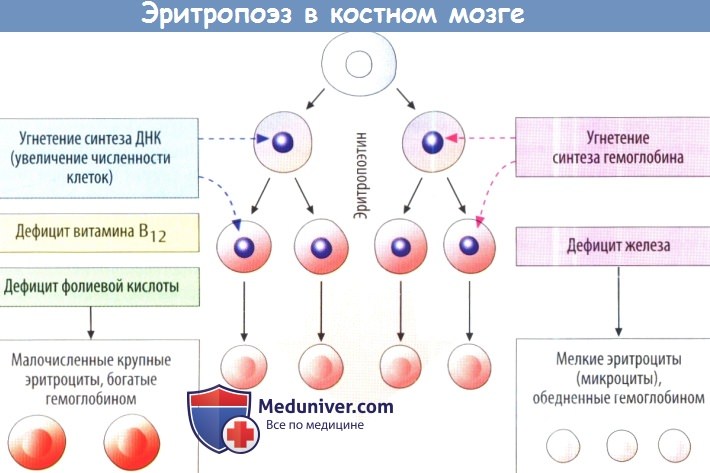
Витамин В12 и фолиевая кислота (витамин В9) необходимы для синтеза нуклеопротеинов в разных тканях организма, созревания и деления ядер эритроидных клеток в кроветворной ткани. При дефиците витаминов В12 и В9 в наиболее интенсивно делящейся ткани организма — эритроидной — раньше, чем в других, возникают нарушения, вызывающие анемию. При дефиците витамина В12 в костном мозге появляются большие ядросодержащие эритроидные клетки — мегалобласты, которые образуют с замедленной скоростью большие эритроциты — мегалоциты с резко укороченным периодом жизни. Замедление поступления эритроцитов в кровь и быстрое их разрушение ведет к анемии. Дефицит витамина В12 возникает в организме при утрате париетальными клетками желудка способности продуцировать «внутренний фактор» — гликопротеин (молекулярная масса 60 000). Фактор связывает витамин В12, поступающий с пищей, и предохраняет его от расщепления пищеварительными ферментами. Эти нарушения возникают при атрофии слизистой оболочки желудка, эпителия двенадцатиперстной кишки, часто наблюдающейся, например, у стариков. И хотя запаса витамина В12 в печени достаточно взрослому человеку на 1—5 лет, постепенное его истощение приводит к заболеванию. Суточная потребность в витамине В,2 — 5 мкг, содержание в плазме крови — 150—450 мкг/л. В кишечнике комплекс гликопротеин—витамин В12 фиксируется специальными рецепторами слизистой оболочки тонкого кишечника, далее витамин поступает в интестинальные клетки, затем в кровь и переносится с помощью особых молекул — транскобаламинов (I, II, III типов). Транскобала-мины I и III типа продуцируются лейкоцитами, II — макрофагами. Поэтому при выраженном лейкоцитозе отмечается гипервитаминоз В12. Витамин В12 содержится в больших количествах в печени, почках, куриных яйцах.
Фолиевая кислота (витамин В9) поддерживает синтез ДНК в клетках костного мозга, благодаря обеспечению этого процесса одним из нуклеоти-дов — диокситимидилатом, образующимся в результате митилирования ди-оксиуридиловой кислоты в присутствии тетрагидрофолата (одной из редуцированных форм фолиевой кислоты). При дефиците витамина В9 в пище у человека уже через 1—6 мес нарушается синтез ДНК и деление эритроидных клеток, ускоряется разрушение эритроцитов, что ведет к анемии. Суточная потребность организма человека в фолиевой кислоте 500—700 мкг. Ее резерв в организме равен 5—10 мг, 1/3 которого находится в печени. Фолиевой кислотой богаты овощи (шпинат), дрожжи, молоко.
1,2,5-дигидроксивитамин D3 и ретиноловая кислота (производное витамина А) участвуют в организме человека в дифференциации кроветворных клеток до их зрелых форм. Витаминами А и D богаты печень трески, тунца, сельдь, коровье молоко и сливочное масло.
Витамин В6 (пиридоксин) является кофактором (т. е. дополнительным фактором активности) фермента — синтетазы 5-аминолевулиновой кислоты (АЛК-синтетазы), участвующей в образовании гема в эритроидных клетках в ткани костного мозга (см. рис. 7.2). Дефицит витамина В6 в организме человека нарушает синтез гемоглобина и вызывает анемию. Витамином В6 богаты зерна злаков, капуста, картофель, молоко.
Витамин С поддерживает основные этапы эритропоэза, способствуя метаболизму фолиевой кислоты в эритробластах. Он участвует в метаболизме железа, повышая как его абсорбцию в желудочно-кишечном тракте, так и мобилизацию депонированного в клетках железа.
Витамин Е (токоферол) и витамин РР защищают фосфатидилэтанол амин мембраны эритроцитов от усиливающего их гемолиз перекисного окисления.
Защиту гемоглобина и мембраны эритроцитов от окисления осуществляет и витамин РР, являющийся одной из составляющих пиридиннуклео-тидов НАД и НАДФ.
Дефицит витамина В2, участвующего в окислительно-восстановительных реакциях, вызывает у человека анемию вследствие замедленного образования эритроцитов в костном мозге.
Источник
Синтез эритроцитов дефицит витамина
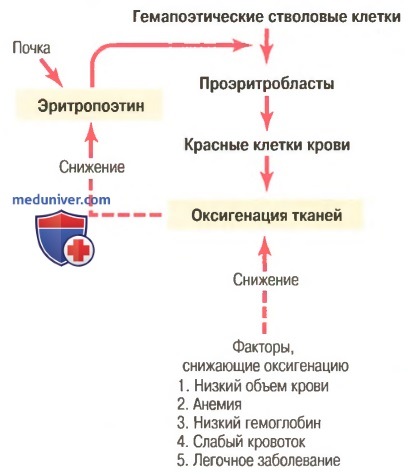
а) Влияние эритропоэтина на эритрогенез. При помещении животного или человека в атмосферу с низким парциальным давлением кислорода эритропоэтин начинает формироваться в течение нескольких минут или часов и достигает максимальной продукции в течение 24 ч. Однако на протяжении примерно 5 сут в циркулирующей крови новые красные клетки практически не появляются.
 Увеличение продукции красных клеток крови, связанное с функцией механизма эритропоэтина при снижении оксигенации тканей
Увеличение продукции красных клеток крови, связанное с функцией механизма эритропоэтина при снижении оксигенации тканей
На основании этого факта, а также результатов других исследований установлено, что важным эффектом эритропоэтина является стимуляция продукции проэритробластов из гемопоэтических стволовых клеток в костном мозге. Кроме того, сразу после формирования проэритробластов эритропоэтин способствует более быстрому, чем в норме, прохождению этих клеток через разные эритробластические стадии, еще больше ускоряя продукцию новых красных клеток крови.
Быстрое образование клеток продолжается до тех пор, пока человек находится в условиях низкого давления кислорода или пока количество эритроцитов не станет достаточным для переноса адекватного количества кислорода к тканям, несмотря на низкое парциальное давление кислорода. В этом случае скорость продукции эритропоэтина снижается до уровня, способного поддерживать необходимое, но не избыточное количество красных клеток крови.
При отсутствии эритропоэтина костный мозг формирует очень мало эритроцитов. С другой стороны, если эритропоэтина синтезируется много, а железо и другие необходимые питательные вещества представлены в изобилии, скорость продукции красных клеток крови может возрасти в 10 и более раз по сравнению с нормой. Следовательно, механизм эритропоэтина для регуляции продукции эритроцитов является очень мощным.
б) Созревание красных клеток крови — потребность в витамине В12 (цианкобаламине) и фолиевой кислоте. В связи с постоянной потребностью в поступлении новых красных клеток в кровь эритропоэтические клетки костного мозга являются одними из наиболее быстрорастущих и размножающихся клеток в организме. Следовательно, их созревание и скорость продукции в значительной степени зависят от состояния питания человека.
Особенно важны для окончательного созревания красных клеток крови два витамина: витамин В12 и фолиевая кислота. Оба витамина необходимы для синтеза ДНК, поскольку каждый из них разным путем участвует в формировании тимидинтрифосфата — одного из важных стандартных блоков ДНК.
Следовательно, недостаток витамина B12 и фолиевой кислоты ведет к синтезу аномальных и уменьшенных молекул ДНК и нарушению созревания ядер и клеточного деления. Более того, эритробластные клетки костного мозга, кроме неспособности быстро размножаться, образуют в основном более крупные красные клетки крови, называемые макроцитами. У таких клеток очень ломкая мембрана, часто неправильная овальная форма вместо обычной формы двояковогнутого диска.
После выхода в циркулирующую кровь эти плохо сформированные клетки способны нормально переносить кислород, но ломкость резко укорачивает их жизнь: до 1/2 или даже 1/3 нормального срока жизни эритроцитов. В связи с этим дефицит витамина В12 или фолиевой кислоты ведет к нарушению созревания в процессе эритропоэза.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Источник
Параграф 120. Биохимия эритроцитов
Составитель текстов – Анисимова Е.С. Авторские права защищены (продавать текст нельзя). Курсив не зубрить.
ПАРАГРАФ 120. См. сначала 6, 22, 26, 27, 32, 59.
3. Биохимия эритроцитов.
6. Классификация гемопротеинов.
5. Гемоглобин и миоглобин, роль глобина. Гемоглобинопатии. Понятие о синтезе гемоглобина.
4. Роль витаминов и железа. Обмен железа.
1. Транспорт эритроцитами О2 и СО2.
2. Значение антиоксидантной системы для эритроцита.
7. Биохимические свойства нейтрофилов.
120. 1. Транспорт кислорода и углекислого газа в крови.
(Газообмен между тканями и эритроцитами и между лёгкими и эритроцитами).
Транспорт кислорода осуществляется от лёгких к тканям эритроцитами, с током крови.
СО2 транспортируется от тканей к лёгким с участием эритроцитов, но не внутри эритроцитов (см. далее).
О2 нужен клеткам в основном для синтеза АТФ, а также для выработки АФК и ряда реакций.
Поэтому нарушение доставки кислорода в ткани приводит к дефициту кислорола (кислородному голоданию, гипоксии), снижению синтеза АТФ и может закончиться смертью.
Кроме дефицита АТФ, при гипоксии возникает АЦИДОЗ из-за накопления ЛАКТАТА (п.32).
Одной из причин нарушения доставки кислорода в ткани является нарушение работы эритроцитов из-за их дефицита (из-за нарушения образования при гематопоэзе или из-за разрушения (гемолиза)) или из-за дефицита гемоглобина.
СО2 в тканях образуется из разных веществ при их распаде (катаболизме): в ЦТК (п.21), ПДГ, при распаде нуклеотидов (п.71).
В лёгких СО2 выдыхается: поступает из крови в дыхательные пути, затем выдыхается.
Кислород при вдохе поступает из окружающей среды в лёгкие, в альвеолах он поступает в кровь и связывается гемоглобином эритроцитов.
Транспорт кислорода эритроцитами осуществляется с помощью белка ГЕМОГЛОБИНА (Hb).
В лёгких гемоглобин связывается с молекулами кислорода, затем с током крови эритроциты поступают в ткани.
Газообмен между тканями и эритроцитами.
1. В тканях кислород отделяется от гемоглобина и переходит из эритроцитов в клетки.
Отделению кислорода от гемоглобина способствует вещество, которое называется 2,3-БИСФОСФО/ГЛИЦЕРАТ и образуется в особой реакции гликолиза, которая протекает в эритроцитах.
Без 2,3-бФГ поступление кислорода в ткани было бы снижено. 2,3-бФГ способствует отделению кислорода от гемоглобину за счёт связывания с молекулами гемоглобина и изменения их конформации (см. п.58). При изменении конформации гемоглобина молекулы кислорода быстрее отделяются от молекул гемоглобина. Используют выражение «снизилось сродство гемоглобина к кислороду», то есть способность гемоглобина связываться с кислородом.
2. При поступлении кислорода в клетку из клетки в эритроцит поступает молекула СО2.
3. В эритроците СО2 вступает в реакцию с молекулой воды, в результате чего образуется угольная кислота – Н2СО3. Реакция катализируется ферментом карбангидраза (карбонатдегидратаза), которому нужен цинк в качестве кофактора (п.4). Реакция обратимая, в тканях равновесии реакции смещено в сторону образования угольной кислоты.
4. Молекула угольной кислоты диссоциирует на Н+ (протон) и анион НСО3 – (бикарбонат).
5. Протон (Н+) связывается с молекулой гемоглобина, а
бикарбонат выходит из эритроцита в кровяное русло.
Способ транспорта бикарбоната через мембрану эритроцита:
выход бикарбоната из эритроцита происходит в обмен на вход в эритроцит иона хлора (Сl – ) = хлорида = хлорид-аниона. То есть в антипорте с хлоридом.
Благодаря этому антипорту заряд мембраны эритроцитов не меняется.
В итоге от тканей в лёгкие транспортируется не молекула СО2 в эритроцитах, а образованные из неё протон и бикарбонат: протон связан с гемоглобином, а бикарбонат в кровяном русле.
Реакции в эритроцитах при их нахождении в тканях:
СО2 + Н2О ; Н2СО3 (угольная кислота) ; Н+ (протон, связывается с Hb) + НСО3 – (бикарбонат,
выходит из эритроцита в антипорте с входящим в эритроцит хлоридом).
Газообмен между лёгкими и эритроцитами.
Когда кровь проходит через лёгкие, в находящихся в крови эритроцитах происходят процессы, противоположные процессам, которые проткали в тканях (см. выше).
1. Бикарбонат входит в эритроцит в обмен на выходящий хлорид (в антипорте с ним).
2. Бикарбонат соединяется (ассоциирует) с протоном, который был связан с гемоглобином,
в результате чего образуется молекула угольной кислоты.
3. Молекула угольной кислоты распадается на молекулу воды и молекулу углекислого газа, которая выходит из эритроцита, а затем выдыхается.
4. Молекулы кислорода поступают в эритроциты и связываются с гемоглобином;
гемоглобин, связанный с кислородом, называется ОКСИ/ГЕМОГЛОБИНОМ.
Реакции в эритроцитах при их нахождении в лёгких:
Н+ (отсоединяется от Hb) + НСО3 – (бикарбонат, входит в эритроцит в антипорте с выходящим из эритроцита хлоридом) ; Н2СО3 (угольная кислота) ; СО2 + Н2О
Таким образом, участие эритроцитов в транспорте газов не требует затрат АТФ.
2. Значение антиоксидантной системы для эритроцита. См. п.27.
Кратко: из молекул кислорода, транспортируемых эритроцитами, могут образовываться активные формы кислорода – АФК. АФК способны разрушать эритроциты, что может привести к кислородному голоданию тканей и смерти – п.22. Чтобы этого не произошло – эритроцитам нужны средства для обезвреживания АФК и предотвращения разрушения эритроцитов – антиокислительная система.
Окисление иона железа гемоглобина кислородом.
Молекулы кислорода в эритроцитах связаны с гемоглобином. В состав гемоглобина входит гемма, в состав которого входит ион железа Fе2+.
Молекула кислорода способна «забирать» один электрона у Fe2+ гемоглобина, превращая его в Fe3+
и превращаясь сама в молекулу кислорода с одним лишним электроном, которая обозначается как •О2 –, называет супероксидрадикалом (СОР) и относится к АФК.
Гемоглобин с Fe3+ вместо Fe2+ называется МЕТгемоглобином и обозначается как MetHb.
Реакция образования метгемоглобина:
Гемоглобин с Fe2+ + О2 ; метгемоглобин с Fe3+ + •О2 – (супероксидрадикал, АФК).
Восстановление метгемоглобина.
Проблема в том, что MetHb не способен транспортировать кислород, поэтому превращение нормального гемоглобина в метгемоглобин нарушает доставку кислорода в ткани и приводит к гипоксии, дефициту АТФ, ацидозу.
Чтобы не было этих негативных последствий образования метгемоглобина, его нужно превращать в гемоглобин. Для этого нужно присоединить электрон к Fe3+, то есть восстановить Fe3+.
Источником электрона для восстановления метгемоглобина является молекула НАДН, образующаяся при гликолизе в эритроцитах (п.32).
(Это одна из причин, по которым эритроцитам нужна глюкоза, которую добавляют в кровь при её хранении).
Фермент, катализирующий присоединение электрона к метгемоглобину за счёт переноса электрона от НАДН, называется МЕТГЕМОГЛОБИН/РЕДУКТАЗОЙ.
Реакция восстановления метгемоглобина:
Метгемоглобин с Fe3+ + НАДН из гликолиза ; гемоглобин с Fe2+ + НАД+.
Из-за того, что ион железа гемоглобина должен всегда быть в состоянии Fe2+,
говорят, что у иона железа гемоглобина постоянная валентность.
А у ионов железа цитохромов валентность переменная – это означает, что их ион железа в процессе функционирования превращается из Fe2+ в Fe3+ и обратно.
Из-за того, что восстановление метгемоглобина происходит внутри эритроцитах, гемоглобин не может транспортировать кислород вне эритроцитов, если попадает в кровяное русло при разрушении эритроцитов при гемолизе.
Перекисное окисление липидов мембран эритроцитов.
Другая проблема образования метгемоглобина в том, что при этом образуется активная форма кислорода – СОР.
АФК вступают в реакции с липидами мембран эритроцитов (с остатками жирных кислот):
АФК «забирают» электроны у липидов
превращая липиды мембран в окисленные липиды мембран (липиды с группами –О-О-Н, которые относятся к органическим перекисям наряду с другими органическими веществами, в которых образовались перекисные группы)
– эти реакции называются ПЕРЕКИСНЫМ ОКИСЛЕНИЕМ ЛИПИДОВ (ПОЛ)
и бывают не только в эритроцитах, но и в других клетках.
ПОЛ эритроцитов приводит к разрушению мембран и эритроцитов в целом, то есть к ГЕМОЛИЗУ.
Гемолиз приводит к таким последствиям, как гипоксия (а она – к дефициту АТФ и ацидозу) и отравление билирубином (см. 118), а затем к смерти.
Восстановление окисленных молекул липидов мембран эритроцитов.
Чтобы гемолиз не произошёл, липиды мембран, окисленные активными формами кислорода, нужно восстанавливать за счёт присоединения к ним электронов.
При восстановлении окисленных липидов перекисные группы (-О-О-Н) липидов превращаются в СПИРТОВЫЕ (-ОН).
Для восстановления липидов нужны источники электронов и катализаторы переноса электронов (ферменты, антиокислительные).
Источником электронов для восстановления липидов является ГЛУТАТИОН (п.27 и 56):
2 молекулы глутатиона отдают по одному атому водорода (и по одному электрону в их составе) для восстановления липидов мембран;
при этом атом серы одной молекулы глутатиона соединяется с атомом серы другой молекулы глутатиона, в результате чего образуется молекула, которую называют окисленной формой глутатиона, которая обозначается так: GS-SG. Молекула глутатиона до отдачи атома водорода обозначается GSH и называется восстановленной формой глутатиона.
Фермент, который катализирует восстановление органических перекисей
за счёт превращения перекисных групп органических перекисей в спиртовые группы, называется ГЛУТАТИОН/ПЕРОКСИДАЗОЙ (ГПО). Приставка «глутатион» связана с тем, что именно глутатитон является источником электронов для восстановления.
Реакция ГПО:
R-O-O-H + GSH + HSG; R-O-H + Н-ОН + GS-SG.
Количество молекул глутатиона в эритроците ограничено. Поэтому окисленный глутатион нужно постоянно превращать в восстановленный: GSSG в (2)GSH. Для восстановления глутатиона нужно расщепить связь между атомами серы окисленной формы глутатиона и присоединить к обоим атомам серы по одному атому водорода.
Источником 2 атомов водорода для восстановления глутатиона является НАДФН, Н+, как и во многих других процессах – см. п.35.
Фермент, который катализирует присоединение водорода к глутатиону (его окисленной форме), называется ГЛУТАТИОН/РЕДУКТАЗОЙ (ГР).
Реакция глутатионредуктазы:
GS-SG + НАДФН, Н+ ; GSH + HSG + НАДФ+.
Количество молекул НАДФН, Н+ в эритроците ограничено. Поэтому НАДФ+ нужно постоянно превращать в НАДФН, Н+:
это превращение происходит в реакциях пентозофосфатного пути, для которого нужен витамин В1.
Вариант ПФП, который протекает в эритроцитах, называется пентозным циклом.
Исходным субстратом ПФП является молекула глюкозы. Это ещё одна причина, по которой эритроциты должны получать глюкозу.
Из-за того, что продукт ПФП (НАДФН) участвует в предотвращении гемолиза –
нарушение ПФП может стать причиной гемолиза.
Причиной нарушения ПФП может стать сниженная активность ферментов ПФП из-за мутации генов.
Встречаются люди со сниженной активностью ферментов ПФП, которые страдают из-за гемолиза.
Гемолиз приводит к анемии. Такие анемии, причиной которой является гемолиз, называются гемолитическими.
Кроме недостаточной активности ферментов ПФП, причиной гемолиза может быть резус –конфликт у новорожденного, поступление в организм гемолитических ядов, серповидно-клеточная анемия и т.д..
3. Биохимия эритроцитов.
У эритроцитах специфический обмен веществ. В них нет органелл. Есть внешняя мембрана, гемоглобин и ряд ферментов, о некоторых из которых уже шла речь.
Из-за отсутствия ядра в эритроцитах нет репликации и транскрипции,
из-за отсутствия митохондрий в эритроцитах нет ;-окисления жирных кислот, ЦТК, ДЦ, синтеза АТФ способом окислительного фосфорилирования.
Нет синтеза и распада гликогена и многих других процессов.
Из процессов обмена веществ в эритроцитах есть уже упоминавшиеся гликолиз и ПФП (вариант «пентозный цикл»), для которых нужна глюкоза.
ПФП нужен эритроцитам для выработки НАДФН для антиокислительной системы.
Гликолиз нужен эритроцитам для получения НАДН для восстановления метгемоглобина,
для получения 2,3-бисфосфо/глицерата для поступления кислорода в ткани (выше),
для получения АТФ:
2 молекул АТФ способом субстратного фосфорилирования (п.23 и 32) в расчёте на одну молекулу глюкозы.
Окончательным метаболитом гликолиза в эритроцитах является ЛАКТАТ, поскольку в них нет митохондрий, в которых лактат мог бы окисляться.
Такой вариант гликолиза (завершающийся образованием лактата) называется анаэробным гликолизом.
В эритроцитах гликолиз является единственным источником АТФ.
АТФ в эритроцитах используется в качестве источника энергии для работы натрий-клиевой-АТФ-азы.
Эритроциты образуются из клеток красного костного мозга, как и все клетки крови,
в ходе гематопоэза (кроветворения): процесса, при котором клетки красного мозга превращаются в определённые клетки крови. То есть в ходе дифференцировки.
Превращение клеток красного мозга в клетки-предшественники эритроцитов происходит под влиянием определённых гормонов, в том числе под влиянием ЭРИТРОПОЭТИНА – гормона, который вырабатывается ПОЧКАМИ;
при тяжёлой почечной недостаточности эритропоэтина вырабатывается недостаточно, что может привести к дефициту эритроцитов и развитии АНЕМИИ – такая анемия (причиной которой является дефицит эритропоэтина) лечится только эритропоэтином;
эритропоэтин получают генно-инженерным способом, как и инсулин.
Предшественники эритроцитов являются клетками, в которых есть все органеллы: митохондрии, ядро и т.д.
Предшественники эритроцитов, которые образуются из клеток красного костного мозга,
называются ЭРИТРОБЛАСТАМИ (бластами часто называются молодые клетки – например, остеобласты).
В эритробластах есть ещё все органеллы, в том числе ядро.
Наличие в эритробластах ядра даёт им возможность синтезировать РНК. В основном синтезируются РНК, нужные для синтеза гемоглобина, а также других белков эритроцитов.
Затем происходит разрушение ядра, в результате чего эритробласты превращаются в ретикулоциты, поскольку в них пока ещё есть ретикулум (ЭПР) и все остальные органеллы, кроме ядра. Наличие в ретикулоцитах ранее синтезированных РНК позволяет им активно синтезировать гемоглобин.
Затем происходит разрушение всех органелл ретикулоцита, в результате чего ретикулоцит превращается в зрелый эритроцит.
4. Роль витаминов и железа (в синтезе гемоглобина и предотвращении анемии). Обмен железа.
Железо человеку нужно для образования в организме гема для гемопротеинов.
Кроме железа, в состав гема входит органическая часть – порфириновое кольцо.
Кроме гема, железо нужно для железо-серных центров дыхательной цепи.
Недостаточное поступление железа в организм может привести к дефициту гемоглобина.
Форма анемии, причиной которой является дефицит железа, называется железо-дефицитной. Есть другие формы анемий, у которых другие причины.
Избыточное содержание железа в организме тоже вредно.
1. Поступление железа в организм с пищей.
Изначально железо поступает в организм с пищей (гранаты, яблоки, икра, мясо и т.д.).
Поэтому неполноценное питание, при котором в рационе мало продуктов, содержащих железо, приводит к анемии. Такая анемия, причиной которой является дефицит в пище железа, называется железо-дефицитной анемией. Она лечится введением в рацион продуктов, содержащих железо.
В сутки в организм с пищей должно поступать 10-15 мг железа.
В мясе железо содержится в составе гема миоглобина и поэтому называется гемовым;
железо мяса двухвалентное, то есть ион железа в состоянии Fe2+.
В большинстве других продуктов железо трёхвалентное (то есть с ионом Fe3+).
2. Всасывание железа в кишечнике.
В кишечнике может усваиваться только двухвалентное железо, поэтому нужно превращать Fe3+ в Fe2+.
Для превращения Fe3+ в Fe2+ нужно присоединить электрон к Fe3+, то есть восстановить Fe3+.
Источником электронов для восстановления трёхвалентного железа в кишечнике является АСКОРБАТ (витамин С).
Поэтому дефицит витамина С в пище приводит к анемии, которая не вылечивается только лишь железом.
В аптеке есть препараты, в которых сочетаются железо и витамин С.
Железо-дефицитная анемия и анемия, обусловленная дефицитом витамина С, похожи тем, что они обе обусловлены дефицитом в пище нужных организму веществ, то есть нарушением питания, то есть это алиментарные анемии.
Из 10-15 грамм железа, которые должны поступать в сутки в организм, всасывается только 1 грамм. Столько же железа организм теряет в сутки (в основном с калом).
3. Транспорт и хранение железа в организме.
Из кишечника железо поступает в кровь, в которой транспортируется в комплексе с белком-транспортёром железа, который называется ТРАНСФЕРРИНОМ.
Трансферрин транспортирует железо в органы, которые выполняют функцию хранилищ (депо) железа: печень и красный костный мозг.
В клетках органов-хранилищ железо хранится в виде комплексов с белком ФЕРРИТИНОМ.
Из органов-депо железа железо по мере необходимости транспортируется кровотоком (в комплексе с трансферрином) во все клетки, которым нужно железо
(в основном для включения железа в состав ферментов, в работе которых участвует железо в качестве кофактора).
4. Использование железа в организме.
В самой печени железо используется для работы ряда белков: цитохромов дыхательной цепи и цитохрома Р 450, оксидаз и оксигеназ и т.д.
В красном костном мозге железо используется для образования гемоглобина при образовании эритроцитов при кроветворении.
При разрушении эритроцитов в крови их гемоглобин поступает в кровяное русло и связывается с белком ГАПТОГЛОБИНОМ (см. п. 39 и 90), который относится к белкам острой фазы.
Большая часть железа при распаде эритроцитов и гемоглобина используется организмом вновь (это называется ре-утилизацией).
Но 1 мг железа в сутки теряется организмом.
Для восполнения потери этого одного миллиграмма нужно 1 мг железа усвоить.
Чтобы организм усвоил 1 мг железа – в пище должно быть 10-15 мг железа.
5. Гемоглобин и миоглобин, роль глобина. Гемоглобинопатии. Понятие о синтезе гемоглобина.
Гемоглобин – белок, который находится в эритроцитах, синтезируется в клетках-предшественниках эритроцитов. Выполняет функцию транспорта кислорода от лёгких в ткани, а также обеспечивает транспорт углекислого газа от тканей в лёгкие (п. 121.1).
Состоит из четырёх глобул-субъединиц (то есть является тетрамером), которые активируют друг друга, в чём проявляется положительная кооперативность. За счёт взаимность активации субъединиц активность гемоглобина в 400 раз выше, чем активность одной субъединицы и чем активность белка миоглобина, который похож на субъединицу гемоглобина, находится в мышцах и выполняет в них функцию запасания кислорода.
Глобин – это название белкой части гемоглобина и миоглобина, то есть полипептидной цепи.
Есть ;-глобиновые цепи и ;-глобиновые цепи, они кодируются разными генами.
Мутации в генах, которые кодируют глобин, приводят к нарушениям в синтезе гемоглобина, которые называются талассемиями и относятся к гемоглобино/патиям.
Синтез гемоглобина.
Синтез гемоглобина происходит в клетках-предшественниках эритроцитов при кроветворении.
Нарушения синтеза гемоглобина называются гемоглобинопатиями и приводят к дефициту гемоглобина и развития анемий.
Слово гемоглобин состоит из двух частей – гем и глобин.
Гемоглобин состоит из двух компонентов: белковой части (глобиновых цепей) и небелковой части (кофактора),
которая называется гемом и состоит из порфиринового кольца (порфирина) и иона железа.
Глобиновые цепи образуют глобулы, в состав одной молекулы гемоглобина входит 4 глобиновых глобулы, которые образуют 4 суъединицы.
Ион железа должен поступить с пищей, усвоиться с помощью витамина С и транспортироваться трансферрином в красный костный мозг.
Порфирин должен синтезироваться в эритроцитах из аминокислоты глицина и метаболита ЦТК сукцинилКоА, в состав которого входит витамин пантотенат.
Глобиновые цепи синтезируются так же, как все полипептидные цепи – в ходе трансляции на рибосомах (п.82).
Причины нарушений образования гемоглобина и их коррекция.
Снабжение железом может нарушиться из-за дефицита железа в организме, из-за дефицита витамина С, из-за повреждения кишечника, из-за дефектов трансферрина.
Дефицит железа и витамина С в пище исправляется употреблением продуктов, которые содержат железо и витамин С; кишечник лечится.
Синтез порфирина может нарушиться из-за дефицитов витаминов ЦТК (в том числе пантотената) и из-за мутаций в генах, кодирующих ферменты синтеза порфирина.
Дефицит витаминов исправляется употреблением продуктов, содержащих витамины ЦТК.
Мутации в генах, кодирующих ферменты синтеза порфирина, приводят к ПОРФИРИЯМ.
Синтез глобиновых цепей может нарушиться (как и синтез любых белков) из-за дефицита в пище веществ, нужных для синтеза белка, а также из-за мутаций в генах, кодирующих глобиновые цепи.
Для синтеза глобиновых цепей нужны 20 аминокислот и РНК. Для синтеза РНК нужны нуклеотиды, для синтеза которых нужен ряд витаминов (п.72): РР, В2, В6, фолат и т.д.
Дефицит 20 аминокислот исправляется употреблением продуктов, содержащих 20 аминокислот в составе полноценных белков: мяса, рыбы, молочных, яиц.
Дефицит витаминов исправляется употребление продуктов, содержащих названные витамины.
Мутации в генах, кодирующих глобиновые цепи, приводят к талассемиям.
Серповидно-клеточная анемия развивается из-за мутации в гене, который кодирует глобиновую цепь.
При этом синтезируется неправильная глобиновая цепь, что приводит к разрушению эритроцитов.
Гемоглобинопатии – это нарушения, связанные с нарушением работы, структуры и синтеза гемоглобина.
К ним относятся талассемии, порфирии.
6. Классификация гемопротеинов.
Гемопротеины – это соединения гема и протеина (ППЦ).
То есть сложные белки (п.4), в состав которых входит гем в качестве небелкового компонета (кофактора), а также белковая часть (апопротеин).
Примеры гемопротеинов в организме:
гемоглобин, миоглобин (п.59),
цитохромы дыхательной цепи (п.22), цитохром Р 450 (п.119),
все оксигеназы и оксидазы и т.д.
Ион железа некоторых гемопротеинов должен менять валентность (например, у цитохромов),
а у некоторых гемопротеинов ион железа должен оставаться всегда Fe2+ (у гемоглобина, миоглобина). –
Гемопротеины с переменной валентностью и с постоянной валентностью.
Источник