Пируватдегидрогеназный комплекс
Пируватдегидрогеназный комплекс (ПДК) — комплекс трех ферментов, который осуществляет окислительное декарбоксилирование пирувата. Продуктами окисления являются углекислый газ, ацетил-КоА, НАДН . Н + .
Содержание
Реакция
Реакция, катализируемая пируватдегидрогеназным комплексом:
| Пируват | ПДК | Ацетил-CoA | |
 |  | ||
| CoA-SH + NAD + | CO2 + NADH + H + | ||
Компоненты
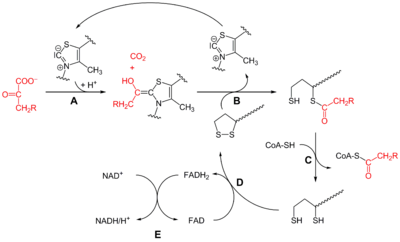

Пируватдегидрогеназный комплекс включает в себя 3 фермента, 3 кофактора (простетическая группа, связанная с апоферментом прочно ковалентно), 2 кофермента (простетическая группа, связанная с апоферментом непрочно нековалентно):
- Е1 — пируватдегидрогеназа декарбоксилирующая.
- Кофактором является активная форма витамина В1 — тиаминпирофосфат.
- Е2 — дигидролипоилацетилтрансфераза.
- Кофактором является витаминоподобное вещество — липоевая кислота, которая присоединив 2 атома водорода может превращаться в дигидролипоил.
- Коферментом является активная форма пантотеновой кислоты — НS-КоА, она принимает ацетильный остаток от липоевой кислоты.
- Е3 — дигидролипоилдегидрогеназа .
- Кофактором является флавинадениндинуклеотид (ФАД) — активная форма витамина В2.
- Коферментом является активная форма витамина PP — никотинамидадениндинуклеотид (НАД + ).
Регуляция
«По механизму обратной связи» ингибируют работу пируватдегидрогеназного комплекса конечные продукты окислительного декарбоксилирования — ацетил-КоА, НАДН . Н + , а также АТФ. Увеличивает активность комплекса пируват.
Также имеется регуляция со стороны гормонов: Инсулин увеличивает активность комплекса, глюкагон — снижает.
Источник
Пируватдегидрогеназный комплекс
Пируватдегидрогеназный комплекс (ПДК) молекулярной массой 6*10 6 д., включает в себя три вида ферментов (Е1-Е3) и пять видов коферментов. При этом 2 кофермента (НАД и HS-КоА) находятся в свободном состоянии и входят в состав комплекса только в момент реакции. Общий вид реакции окислительного декарбоксилирования пирувата:
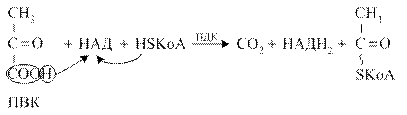
Ферменты пируватдегидрогеназного комплекса
Е1 – пируватдегидрогеназа декарбоксилирующая;
Е2 – дигидролипоилацетилтрансфераза (трансацетилаза);
Коферменты пируватдегидрогеназного комплекса
- Тиаминдифосфат (ТДФ, ТПФ), содержащий витамин В1, кофактор пируватдегидрогеназы.
- Липоевая кислота, кофактор трансацетилазы.
- Кофермент ФАД, содержащий витамин В2, кофактор дегидрогеназы дигидролипоевой кислоты.
- Кофермент НАД, содержащий витамин РР.
- Кофермент НS-КоА, содержащий аденин, рибозу, два остатка фосфорной кислоты, пантотеновую кислоту (витамин В3).
Окислительное декарбоксилирование ПВК протекает в несколько стадий, в процессе которых двухуглеродный фрагмент, образующийся из ПВК, переносится на липоевую кислоту, а затем на HS-КоА.
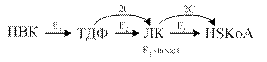
Витамины, входящие в состав пируватдегидрогеназного комплекса
В состав ПДК входит пять витаминов (РР. В2, липоевая кислота, В1, пантотеновая кислота).
Липоевая кислота
Липоевая кислота — витаминоподобное вещество, представляет собой восьмиуглеродную жирную кислоту с двумя -SH группами. Биологическая роль:является коферментом ПДК, участвует в окислении α — кетокислот.
Пантотеновая кислота, витамин В3
Пантотеновая кислота является витамином, который, в свою очередь, включает β — аланин и производное масляной кислоты. Она распространена в животных и растительных продуктах. Суточная потребность в пантотеновой кислоте составляет до 10 мг. Биологическая роль: входит в состав НS- КоА и участвует в окислительном декарбоксилировании α — кетокислот, участвует в активации жирных кислот. Авитаминоз проявляется дерматитом, депигментацией волос, поражением нервной системы.
Витамин В1, тиамин, антиневритный витамин
Витамин В1 включает в свой состав пиримидиновое кольцо, содержит аминогруппу. Суточная потребность в нём составляет 2 мг. Тиамин содержится в злаках, дрожжах. Биологическая роль: входит в состав кофермента ТДФ и участвует в окислительном декарбоксилировании α — кетокислот, а также является коферментом транскетолазной реакции в пентозофосфатном пути окисления глюкозы. Авитаминоз проявляется полиневритами (болезнь бери-бери).
Регуляция пируватдегидрогеназного комплекса осуществляется путём фосфолирирования — дефосфолирирования пируватдегидрогеназы
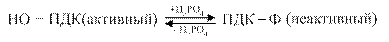
Активаторами ПДК служат АДФ и НАД окисленный. Ингибиторами этого комплекса являются АТФ и НАДН2..
5.1.2. Биологическая роль окислительного декарбоксилирования пирувата Биологическое значение процесса окислительного декарбоксилирования пировиноградной кислоты заключается в том, что оно является важным этапом катаболизма, позволяющим включаться в цикл Кребса тем веществам, при распаде которых образуется ПВК. Образовавшаяся молекула НАДН2 окисляется в длинной дыхательной цепи с образованием 3-х молекул АТФ. Окислительное декарбоксилирование пирувата протекает внутри митохондрий.
Дата добавления: 2016-09-26 ; просмотров: 9036 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Источник
1. Строение пируватдегидрогеназного комплекса
Процесс окислительного декарбоксилирования пирувата катализирует сложнооргани-зованный пируватдегидрогеназный комплекс. В пируватдегидрогеназный комплекс (ПДК) входят 3 фермента: пируватдекарбоксилаза (Е1), дигидролипоилтрансацетилаза (Е2) и дигидролипоилдегидрогеназа (Е3), а также 5 коферментов: тиаминдифосфат (ТДФ), липоевая кислота,. FAD, NAD + и КоА. Кроме того, в состав комплекса входят регуляторные субъединицы: протеинкиназа и фосфопротеинфосфатаза (табл. 6-5).
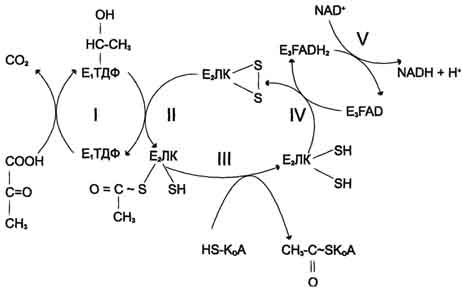
Рис. 6-21. Последовательность реакций, катализируемых ПДК. I — Е1 катализирует декарбоксилирование пирувата и перенос С2-фрагмента на ТДФ; II — Е2 катализирует окисление гидроксиэтильной группы и перенос С2-фрагмента на липоевую кислоту (ЛК); III — ацетилированная дигидролипоилтрансацетилаза взаимодействует с КоА с образованием восстановленной формы липоевой кислоты и ацетил-КоА; IV — окисленная форма трансацетилазы регенерируется при умастии E3;V — окисленная форма Е3 регенерируется при участии NAD + .
Таблица 6-5. Пируватдегидрогеназный комплекс (ПДК) млекопитающих
1. Пируватдекарбоксилаза (пируватдегидрогеназа)
120 (30 тетрамеров)
Липоевая кислота (ЛК)
Все эти ферменты и коферменты объединены в мультиферментную систему, содержащую разные количества каждого из ферментов и имеющую молекулярную массу более 6×10 6 .
В центре комплекса располагается дигидролипоилтрансацетилаза (Е2), образуя его ядро. К дигидролипоилтрансацетилазе присоединены молекулы: пируватдекарбоксилазы (Е1) и дигидролипоилдегидрогеназы (Е3).
Пируватдекарбоксилаза содержит прочно связанный с белковой частью ТДФ, а дигидроли-поилдегидрогеназа — FAD.
Липоиллизиновые группы центрального фермента (Е2) функционируют как поворотные «кронштейны», переносящие атомы водорода и ацетильные группы от одной ферментной молекулы комплекса к другой.
2. Окислительное декарбоксилирование пирувата
Превращение пирувата в ацетил-КоА включает 5 стадий (рис. 6-21).
Стадия I. На этой стадии пируват соединяется с ТДФ в составе Е1 и подвергается декарбоксилированию.
Пируват + Е1-ТДФ → Гидроксиэтил-ТДФ + CO2.
В результате этой реакции образуется производное ТДФ с гидроксиэтильной группой при тиазоловом кольце .
Стадия П. Дигидролипоилтрансацетилаза (Е2) катализирует перенос атома водорода и ацетильной группы от ТДФ на окисленную форму липоиллизиновых групп с образованием ацетилтиоэфира липоевой кислоты .
Стадия III. На стадии III КоА взаимодействует с ацетильным производным Е2, в результате чего образуются ацетил-КоА и полностью восстановленный липоильный остаток, простетическая группа Е2 .
Стадия IV. На стадии IV дигидролипоилде-гидрогеназа (Е3) катализирует перенос атомов водорода от восстановленных липоильных групп на FAD — простетическую группу фермента Е3.
Стадия V . На стадии V восстановленный FADH2 передаёт водород на NAD + с образованием NADH.
Пируватдегидрогеназный комплекс характеризуется большим отрицательным окислительно-восстановительным потенциалом, который обеспечивает наряду с восстановлением кофермента (NADH) образование высокоэнергетической тиоэфирной связи в ацетил-КоА.
Структурное объединение 3 видов ферментов создаёт возможности для координации отдельных этапов сложной ферментативной реакции. Все промежуточные продукты реакции окислительного декарбоксилирования пирувата прочно связаны с комплексом, что увеличивает суммарную скорость процесса и сводит к минимуму побочные реакции.
Пируватдегидрогеназный комплекс, как и все белки, участвующие в реакциях ЦТК, кодируется ядерной ДНК. Транспорт субъединиц ПДК в митохондрии происходит сложным путём за счёт энергии АТФ или трансмембранного электрохимического потенциала при участии белков теплового шока, или шаперонов (см. раздел 1), предотвращающих их преждевременный фолдинг до поступления в митохон-дриальный матрикс или внутреннюю мембрану митохондрий.
Источник
1. Строение пируватдегидрогеназного комплекса
Процесс окислительного декарбоксилирования пирувата катализирует сложноорганизованный пируватдегидрогеназный комплекс. В пируватдегидрогеназный комплекс (ПДК) входят 3 фермента: пируватдекарбоксилаза (Е1), дигидролипоилтрансацетилаза (Е2) и дигидролипоилдегидрогеназа (Е3), а также 5 коферментов: тиаминдифосфат (ТДФ), липоевая кислота,. FAD, NAD + и КоА. Кроме того, в состав комплекса входят регуляторные субъединицы: протеинкиназа и фосфопротеинфосфатаза .
Пируватдегидрогеназный комплекс (ПДК) млекопитающих
1. Пируватдекарбоксилаза (пируватдегидрогеназа)
120 (30 тетрамеров)
Липоевая кислота (ЛК)
Все эти ферменты и коферменты объединены в мультиферментную систему, содержащую разные количества каждого из ферментов и имеющую молекулярную массу более 6×10 6 .
В центре комплекса располагается дигидролипоилтрансацетилаза (Е2), образуя его ядро. К дигидролипоилтрансацетилазе присоединены молекулы: пируватдекарбоксилазы (Е1) и дигидролипоилдегидрогеназы (Е3).
Пируватдекарбоксилаза содержит прочно связанный с белковой частью ТДФ, а дигидроли-поилдегидрогеназа — FAD.
Липоиллизиновые группы центрального фермента (Е2) функционируют как поворотные «кронштейны», переносящие атомы водорода и ацетильные группы от одной ферментной молекулы комплекса к другой.
Окислительное декарбоксилирование пирувата
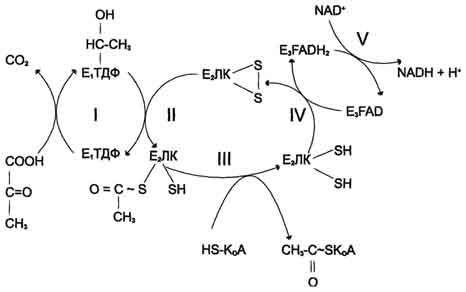
Рис. 6-21. Последовательность реакций, катализируемых ПДК. I — Е1 катализирует декарбоксилирование пирувата и перенос С2-фрагмента на ТДФ; II — Е2 катализирует окисление гидроксиэтильной группы и перенос С2-фрагмента на липоевую кислоту (ЛК); III — ацетилированная дигидролипоилтрансацетилаза взаимодействует с КоА с образованием восстановленной формы липоевой кислоты и ацетил-КоА; IV — окисленная форма трансацетилазы регенерируется при участии E3;V — окисленная форма Е3 регенерируется при участии NAD + .
Пируватдегидрогеназный комплекс характеризуется большим отрицательным окислительно-восстановительным потенциалом, который обеспечивает наряду с восстановлением кофермента (NADH) образование высокоэнергетической тиоэфирной связи в ацетил-КоА.
Структурное объединение 3 видов ферментов создаёт возможности для координации отдельных этапов сложной ферментативной реакции. Все промежуточные продукты реакции окислительного декарбоксилирования пирувата прочно связаны с комплексом, что увеличивает суммарную скорость процесса и сводит к минимуму побочные реакции.
Пируватдегидрогеназный комплекс, как и все белки, участвующие в реакциях ЦТК, кодируется ядерной ДНК. Транспорт субъединиц ПДК в митохондрии происходит сложным путём за счёт энергии АТФ или трансмембранного электрохимического потенциала при участии белков теплового шока, или шаперонов (см. раздел 1), предотвращающих их преждевременный фолдинг до поступления в митохондриальный матрикс или внутреннюю мембрану митохондрий.
Источник
Комплекс пируватдегидрогеназы — Pyruvate dehydrogenase complex

Комплекс пируватдегидрогеназы ( PDC ) — это комплекс из трех ферментов, который превращает пируват в ацетил-КоА с помощью процесса, называемого декарбоксилированием пирувата . Затем ацетил-КоА можно использовать в цикле лимонной кислоты для осуществления клеточного дыхания , и этот комплекс связывает метаболический путь гликолиза с циклом лимонной кислоты . Декарбоксилирование пирувата также известно как «реакция пируватдегидрогеназы», поскольку оно также включает окисление пирувата.
Этот мультиферментный комплекс структурно и функционально связан с мультиферментными комплексами оксоглутаратдегидрогеназы и разветвленной цепи оксокислот дегидрогеназы .
СОДЕРЖАНИЕ
Реакция
Реакция, катализируемая комплексом пируватдегидрогеназы:
| пируват | пируватдегидрогеназный комплекс | ацетил-КоА | |
 |  | ||
| CoA-SH + NAD + | СО 2 + НАДН + Н + | ||
Состав
Пируватдегидрогеназа (E1)

Субъединица E1, называемая субъединицей пируватдегидрогеназы , имеет структуру, состоящую из двух цепей («ɑ» и «ꞵ» цепи). Ион магния образует 4-координатный комплекс с тремя полярными аминокислотными остатками (Asp, Asn и Tyr), расположенными на альфа-цепи, и кофактором тиаминдифосфата (TPP), непосредственно участвующим в декарбоксилировании пирувата .
Дигидролипоил трансацетилаза (E2)
Субъединица E2, или дигидролипоилацетилтрансфераза, как для прокариот, так и для эукариот, обычно состоит из трех доменов. N-концевой домен (липоильный домен) состоит из 1-3 липоильных групп, приблизительно из 80 аминокислот каждая. Домен связывания периферической субъединицы (PBSD) служит селективным сайтом связывания для других доменов субъединиц E1 и E3. Наконец, С-концевой (каталитический) домен катализирует перенос ацетильных групп и синтез ацетил-КоА.
Дигидролипоилдегидрогеназа (E3)

Субъединица E3, называемая ферментом дигидролипоилдегидрогеназа , характеризуется как гомодимерный белок, в котором два остатка цистеина, участвующие в дисульфидных связях, и кофактор FAD в активном центре облегчают ее основное назначение в качестве катализатора окисления. Один пример структуры E3, обнаруженный в Pseudomonas putida , сформирован таким образом, что каждая индивидуальная субъединица гомодимера содержит два связывающих домена, ответственных за связывание с FAD и NAD, а также центральный домен и интерфейсный домен.
Белок, связывающий дигидролипоилдегидрогеназу (E3BP)
Вспомогательный белок, уникальный для большинства эукариот, — это связывающий белок E3 (E3BP), который служит для связывания субъединицы E3 с комплексом PDC. В случае человеческого E3BP гидрофобные остатки пролина и лейцина в BP взаимодействуют с поверхностным сайтом узнавания, образованным связыванием двух идентичных мономеров E3.
Механизм
| Ферменты | Аббревиатура | Кофакторы | # субъединицы прокариот | # субъединицы эукариоты |
|---|---|---|---|---|
| пируватдегидрогеназа ( EC 1.2.4.1 ) | E1 | TPP (тиаминпирофосфат) | 24 | 30 |
| дигидролипоил трансацетилаза ( EC 2.3.1.12 ) | E2 | липоат- кофермент А | 24 | 60 |
| дигидролипоилдегидрогеназа ( EC 1.8.1.4 ) | E3 | FAD NAD + | 12 | 12 |
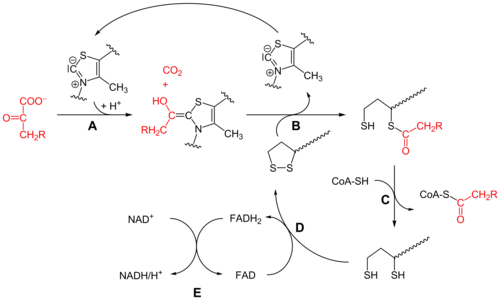
Пируватдегидрогеназа (E1)
Первоначально пируват и тиаминпирофосфат (TPP или витамин B 1 ) связываются субъединицами пируватдегидрогеназы . Тиазолиевое кольцо ТЭС находится в цвиттерионных форме, и анионные C2 углерод выполняет нуклеофильную атаку на С2 (кетон) карбонил пируваты. Полученный гемитиоацеталь подвергается декарбоксилированию с образованием эквивалента ацильного аниона (см. Химию циангидрина или альдегид-дитиан умполунг , а также конденсацию бензоина ). Этот анион атакует S1 окисленных липоатов, которые присоединены к остатку лизина . В S N 2-подобном механизме раскрытия кольца S2 замещается как сульфидный или сульфгидрильный фрагмент. Последующий коллапс тетраэдрического гемитиоацеталя выбрасывает тиазол, высвобождая кофактор TPP и генерируя тиоацетат на S1 липоата. Процесс, катализируемый E1, является лимитирующей стадией всего комплекса пируватдегидрогеназы.
Дигидролипоил трансацетилаза (E2)
На данный момент, липоаты-тиоэфир функциональность транслоцируется в dihydrolipoyl transacetylase активного сайта (Е2), где реакция transacylation передает ацетил из «поворотного кронштейна» от lipoyl к тиолу коэнзим А . Это производит ацетил-КоА , который высвобождается из ферментного комплекса и впоследствии входит в цикл лимонной кислоты . E2 также может быть известен как липоамидредуктаза-трансацетилаза.
Дигидролипоилдегидрогеназа (E3)
Dihydrolipoate , по- прежнему связан с остатком лизина комплекса, затем мигрирует к dihydrolipoyl дегидрогеназы активного сайта (Е3), где он подвергается Flavin опосредованного окисления, идентичного по химии к дисульфидной изомеразе . Во-первых, FAD окисляет дигидролипоат обратно до состояния покоя липоата, производя FADH 2 . Затем кофактор NAD + окисляет FADH 2 обратно до состояния покоя FAD, производя NADH.
Структурные различия между видами
PDC — это большой комплекс, состоящий из нескольких копий 3 или 4 субъединиц в зависимости от вида.
Грамотрицательные бактерии
У грамотрицательных бактерий, например Escherichia coli , PDC состоит из центрального кубического ядра, состоящего из 24 молекул дигидролипоилтрансацетилазы (E2). До 24 копий пируватдегидрогеназы (E1) и 12 молекул дигидролипоилдегидрогеназы (E3) связываются с внешней стороной ядра E2.
Грамположительные бактерии и эукариоты
Напротив, у грамположительных бактерий (например, Bacillus stearothermophilus ) и эукариот центральное ядро PDC содержит 60 молекул E2, расположенных в виде икосаэдра. Это «ядро» субъединицы E2 координируется с 30 субъединицами E1 и 12 копиями E3.
Эукариоты также содержат 12 копий дополнительного корового белка, связывающего белка E3 (E3BP), который связывает субъединицы E3 с ядром E2. Точное местонахождение E3BP не совсем понятно. Крио-электронная микроскопия установила, что E3BP связывается с каждой из граней икосаэдра у дрожжей. Однако было высказано предположение, что он заменяет эквивалентное количество молекул E2 в ядре PDC крупного рогатого скота.
До 60 молекул E1 или E3 могут связываться с ядром E2 грамположительных бактерий — связывание является взаимоисключающим. У эукариот E1 специфически связывается с E2, тогда как E3 ассоциируется с E3BP. Считается, что присутствует до 30 ферментов E1 и 6 E3, хотя точное количество молекул может варьироваться in vivo и часто отражает метаболические потребности рассматриваемой ткани.
Регулирование
Пируватдегидрогеназа ингибируется, когда увеличивается одно или несколько из трех следующих соотношений: АТФ / АДФ , НАДН / НАД + и ацетил-КоА / КоА .
У эукариот PDC строго регулируется собственной специфической киназой пируватдегидрогеназы (PDK) и фосфатазой пируватдегидрогеназы (PDP), дезактивируя и активируя ее соответственно.
- PDK фосфорилирует три специфических остатка серина на E1 с различным сродством. Фосфорилирование любого из них (с использованием АТФ ) делает E1 (и, как следствие, весь комплекс) неактивным.
- Дефосфорилирование E1 PDP восстанавливает комплексную активность.
Продукты реакции действуют как аллостерические ингибиторы PDC, поскольку они активируют PDK. Субстраты, в свою очередь, ингибируют PDK, реактивируя PDC.
Во время голодания количество PDK увеличивается в большинстве тканей, включая скелетные мышцы , за счет увеличения транскрипции генов . В тех же условиях количество PDP уменьшается. Результирующее ингибирование PDC предотвращает катаболизм глюкозы и предшественников глюконеогенеза в мышцах и других тканях . Метаболизм смещается в сторону использования жира , в то время как распад мышечного белка для обеспечения предшественников глюконеогенеза сводится к минимуму, а доступная глюкоза сохраняется для использования мозгом .
Ионы кальция играют роль в регуляции PDC в мышечной ткани, потому что они активируют PDP, стимулируя гликолиз при его высвобождении в цитозоль — во время сокращения мышц . Некоторые продукты этой транскрипции высвобождают H2 в мышцы. Со временем это может привести к распаду ионов кальция.
Локализация декарбоксилирования пирувата
В эукариотических клетках декарбоксилирование пирувата происходит внутри митохондриального матрикса после транспорта субстрата, пирувата, из цитозоля . Транспорт пирувата в митохондрии осуществляется через транспортный белок пируваттранслоказу. Пируваттранслоказа транспортирует пируват симпортом с протоном и, следовательно, является активным , потребляя энергию . Альтернативные источники говорят, что «транспорт пирувата через внешнюю митохондриальную мембрану, по-видимому, легко осуществляется через большие неселективные каналы, такие как потенциал-зависимый анион каналы , которые обеспечивают «пассивную диффузию» и транспорт через внутреннюю митохондриальную мембрану, опосредуются митохондриальным переносчиком пирувата 1 (MPC1) и митохондриальным переносчиком пирувата 2 (MPC2).
При попадании в митохондрии пируват декарбоксилируется с образованием ацетил-КоА. Эта необратимая реакция улавливает ацетил-КоА в митохондриях (ацетил-КоА может переноситься из митохондриального матрикса только в условиях высокого содержания оксалоацетата через цитратный челнок, промежуточное соединение ТЦА, которое обычно мало). Углекислый газ, образующийся в результате этой реакции, неполярный и небольшой, он может диффундировать из митохондрий и из клетки.
У прокариот , у которых нет митохондрий, эта реакция либо осуществляется в цитозоле, либо не происходит вовсе.
Эволюционная история
Было обнаружено, что фермент пируватдегидрогеназа, обнаруженный в митохондриях эукариотических клеток, очень похож на фермент из Geobacillus stearothermophilus , который является разновидностью грамположительных бактерий . Несмотря на сходство комплекса пируватдегидрогеназы с грамположительными бактериями, мало сходства с таковым у грамотрицательных бактерий . Сходство четвертичных структур между пируватдегидрогеназой и ферментами у грамположительных бактерий указывает на общую эволюционную историю, которая отличается от эволюционной истории соответствующих ферментов, обнаруженных у грамотрицательных бактерий. Через эндосимбиотическое событие пируватдегидрогеназа, обнаруженная в митохондриях эукариот, указывает на наследственные связи, восходящие к грамположительным бактериям. Комплексы пируватдегидрогеназы имеют много общего с 2-оксокислотой дегидрогеназы с разветвленной цепью (BCOADH), особенно в их субстратной специфичности для альфа-кетокислот. В частности, BCOADH катализирует разложение аминокислот, и эти ферменты были преобладающими в те периоды, когда на доисторической земле преобладала среда, богатая аминокислотами. Субъединица E2 из пируватдегидрогеназы произошла от гена E2, обнаруженного в BCOADH, в то время как оба фермента содержат идентичные субъединицы E3 из-за присутствия только одного гена E3. Поскольку субъединицы E1 обладают отличительной специфичностью к конкретным субстратам, субъединицы E1 пируватдегидрогеназы и BCOADH различаются, но имеют генетическое сходство. Грамположительные бактерии и цианобактерии, которые позже дадут начало митохондриям и хлоропластам, обнаруженным в эукариотических клетках, сохранили субъединицы E1, которые генетически связаны с субъединицами, обнаруженными в ферментах BCOADH.
Клиническая значимость
Дефицит пируватдегидрогеназы (PCDC) может быть результатом мутаций любого из ферментов или кофакторов. Его первичный клинический признак — лактоацидоз . Такие мутации PCDC, приводящие к последующему дефициту продукции NAD и FAD, препятствуют процессам окислительного фосфорилирования, которые играют ключевую роль в аэробном дыхании. Таким образом, ацетил-КоА вместо этого восстанавливается с помощью анаэробных механизмов до других молекул, таких как лактат, что приводит к избытку лактата в организме и связанным с ним неврологическим патологиям.
Хотя дефицит пируватдегидрогеназы встречается редко, существует множество различных генов, когда они мутированы или нефункциональны, которые могут вызывать этот дефицит. Во-первых, субъединица E1 пируватдегидрогеназы содержит четыре разных субъединицы: две альфа-субъединицы, обозначенные как E1-альфа, и две бета-субъединицы, обозначенные как E1-beta. Ген PDHA1, обнаруженный в субъединицах E1-альфа, при мутации вызывает 80% случаев дефицита пируватдегидрогеназы, поскольку эта мутация сокращает белок E1-альфа. Снижение функционального E1 альфа препятствует достаточному связыванию пируватдегидрогеназы с пируватом, тем самым снижая активность всего комплекса. Мутация гена PDHB, обнаруженного в бета-субъединице E1 комплекса, также приводит к дефициту пируватдегидрогеназы. Аналогичным образом, были обнаружены мутации, обнаруженные в других субъединицах комплекса, такие как ген DLAT, обнаруженный в субъединице E2, ген PDHX, обнаруженный в субъединице E3, а также мутация в гене фосфатазы пируватдегидрогеназы, известная как PDP1, все были отслежены. обратно к дефициту пируватдегидрогеназы, в то время как их конкретный вклад в состояние болезни неизвестен.
Источник