Окислительное декарбоксилирование пировиноградной кислоты
Окисление пирувата до ацетил-КоА происходит при участии ряда ферментов и коферментов, объединенных структурно в мультиферментную систему, получившую название «пируватдегидрогеназный комплекс».
На I стадии этого процесса пируват (рис. 10.8) теряет свою карбоксильную группу в результате взаимодействия с тиаминпирофосфатом (ТПФ) в составе активного центра фермента пируватдегидрогеназы (E1). На II стадии оксиэтильная группа комплекса E1–ТПФ–СНОН–СН3 окисляется с образованием ацетильной группы, которая одновременно переносится на амид липоевой кислоты (кофермент), связанной с ферментом дигидроли-поилацетилтрансферазой (Е2). Этот фермент катализирует III стадию – перенос ацетильной группы на коэнзим КоА (HS-KoA) с образованием конечного продукта ацетил-КоА, который является высокоэнергетическим (макроэргическим) соединением.
На IV стадии регенерируется окисленная форма липоамида из восстановленного комплекса дигидролипоамид–Е2. При участии фермента дигидролипоилдегидрогеназы (Е3) осуществляется перенос атомов водорода от восстановленных сульфгидрильных групп дигидролипоамида на ФАД, который выполняет роль простетической группы данного фермента и прочно с ним связан. На V стадии восстановленный ФАДН2 дигидро-липоилдегидрогеназы передает водород на кофермент НАД с образованием НАДН + Н + .
Процесс окислительного декарбоксилирования пирувата происходит в матриксе митохондрий. В нем принимают участие (в составе сложного мультиферментного комплекса) 3 фермента (пируватдегидрогеназа, ди-гидролипоилацетилтрансфераза, дигидролипоилдегидрогеназа) и 5 кофер-ментов (ТПФ, амид липоевой кислоты, коэнзим А, ФАД и НАД), из которых три относительно прочно связаны с ферментами (ТПФ-E1, ли-поамид-Е2 и ФАД-Е3), а два – легко диссоциируют (HS-KoA и НАД).
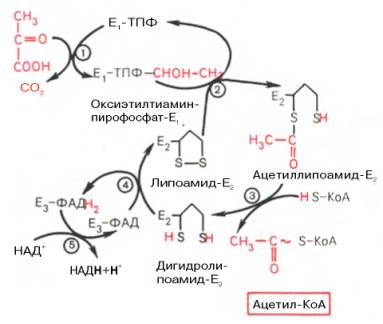
Рис. 10.8. Механизм действия пируватдегидрогеназного комплекса.
Е1 — пируватдегидрогеназа; Е2 — ди-гидролипоилацетилтрансфсраза; Е3 -дигидролипоилдегидрогеназа; цифры в кружках обозначают стадии процесса.
Все эти ферменты, имеющие субъединичное строение, и коферменты организованы в единый комплекс. Поэтому промежуточные продукты способны быстро взаимодействовать друг с другом. Показано, что составляющие комплекс полипептидные цепи субъединиц дигидролипоил-ацетилтрансферазы составляют как бы ядро комплекса, вокруг которого расположены пируватдегидрогеназа и дигидролипоилдегидрогеназа. Принято считать, что нативный ферментный комплекс образуется путем самосборки.
Суммарную реакцию, катализируемую пируватдегидрогеназным комплексом, можно представить следующим образом:
Пируват + НАД + + HS-KoA –> Ацетил-КоА + НАДН + Н + + СO2.
Реакция сопровождается значительным уменьшением стандартной свободной энергии и практически необратима.
Образовавшийся в процессе окислительного декарбоксилирования аце-тил-КоА подвергается дальнейшему окислению с образованием СО2 и Н2О. Полное окисление ацетил-КоА происходит в цикле трикарбоновых кислот (цикл Кребса). Этот процесс, так же как окислительное декарбо-ксилирование пирувата, происходит в митохондриях клеток.
Источник
ОКИСЛИТЕЛЬНОЕ ДЕКАРБОКСИЛИРОВАНИЕ ПВК
Окислительное декарбоксилирование пировиноградной кислоты — многостадийный процесс, который катализирует пируватдегидрогеназный комплекс — митохондриальный мультиферментный комплекс, соединенный с внутренней мембраной со стороны матрикса. ПВК поступает к комплексу из матрикса, и туда же освобождаются продукты реакции.
Пируватдегидрогеназный комплекс включает три фермента (пируватдегидрогеназа (Е1), ацетилтрансфераза (Е2), дегидрогеназа дигидролипоевой кислоты (Е3)) и пять коферментов (НАД + , ФАД, тиаминпирофосфат, липоевая кислота, кофермент А (рис. 36)).
Тиаминпирофосфат связан с пируватдегидрогеназой (Е1), липоевая кислота — с ацетилтрансферазой (Е2), ФАД — с дегидрогеназой дигидролипоевой кислоты (Е3). Кофермент А и НАД + находятся в свободно растворенном состоянии.
В состав пируватдегидрогеназного комплекса входит примерно по три десятка молекул Е1 и Е2 и 10 молекул Е3. Комплекс работает как конвейер: промежуточные продукты не освобождаются в раствор, а передаются от фермента к ферменту.
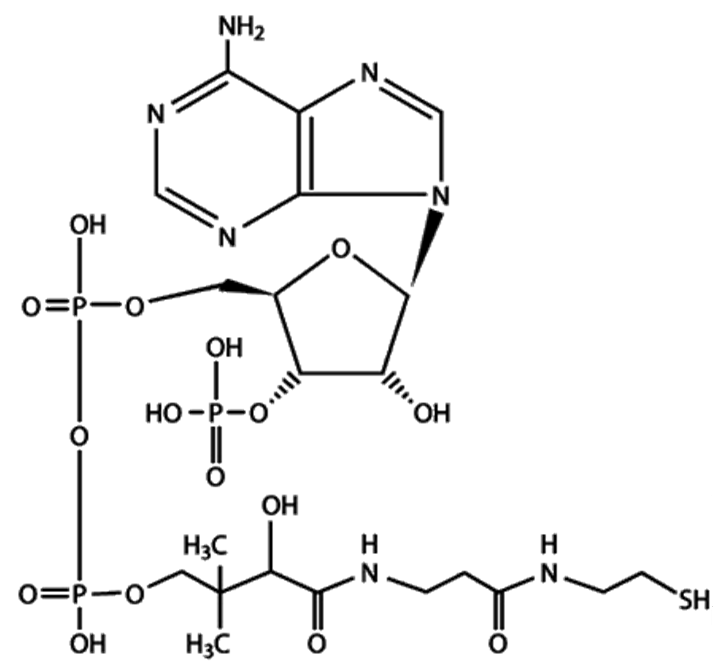 а а | 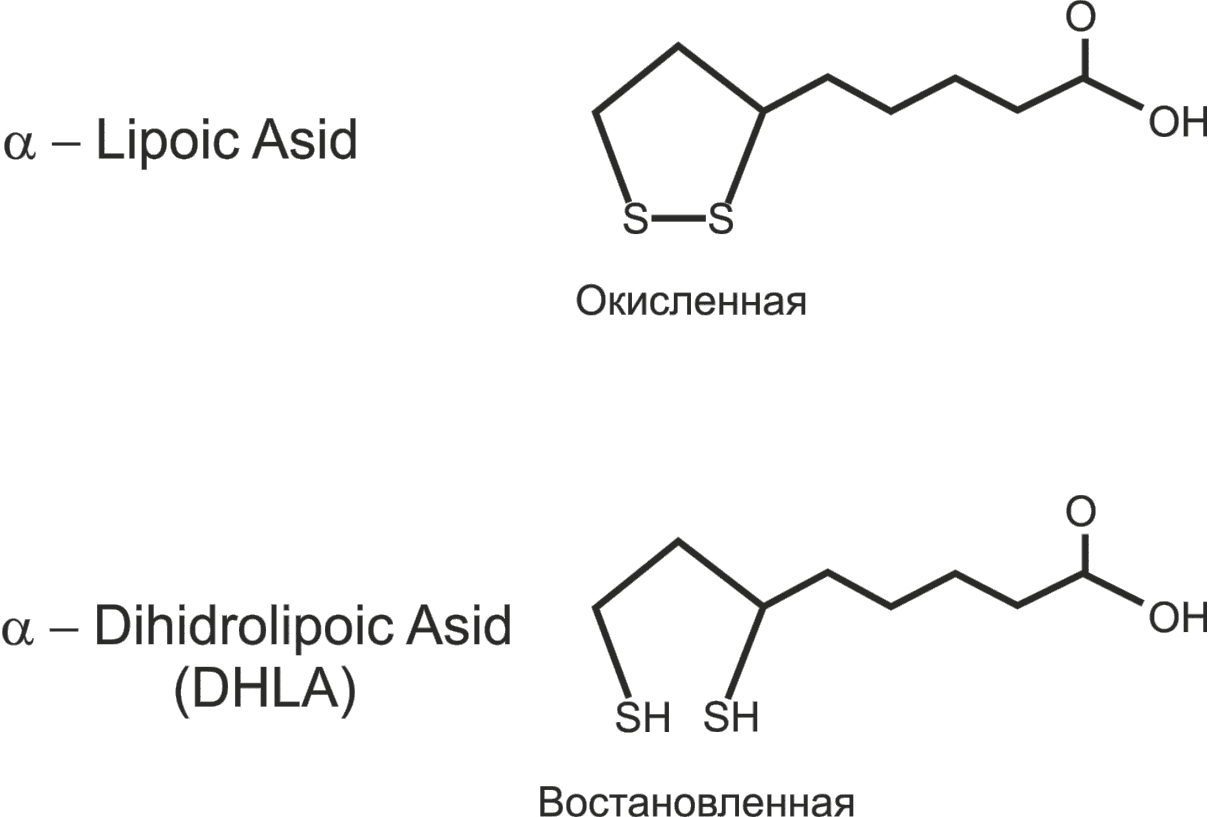 б б |
| Рис. 36. Структура кофермента А (а) и липоевой кислоты (б) |
Первую реакцию (рис. 37) катализирует Е1, субстратами являются ПВК и дегидролипоевая кислота, являющаяся простетической группой Е2. От ПВК отщепляется карбоксильная группа и образуется СО2, а ацетильный остаток соединяется с атомом серы липоевой кислоты в составе ацетилтрансферазы. Образуется ацетиллипоат-Е2.
Во второй реакцииацетилтрансфераза (Е2) катализирует перенос ацетильного остатка, соединенного с его простетической группой, на коэнзим А. Продукты этой реакции — дигидролипоевая кислота в составе Е2 и ацетил-KоА.
В третьей реакциипроисходит дегидрирование дигидролипоевой кислоты в составе ацетилтрансферазы при воздействии фермента Е3 (дегидрогеназа дигидролипоевой кислоты), содержащего ФАД. ФАД передает водород на НАД + . Образуются НАДН+Н + и дегидролипоевая кислота в составе Е2. Последний фермент снова вступает в окислительное декарбоксилирование ПВК.
Ацетил-KоА (продукт второй реакции) затем окисляется в цикле Кребса. Водород с НАДН (продукт третьей реакции) поступает в дыхательную цепь, где образуется АТФ.
Энергетический выход окислительного декарбоксилирования пирувата – 3 АТФ.
Суммарное уравнение процесса:
СH3-CО-COOH + HS-KoA + НАД + ® СH3-CО-S-KoA + НАДН + Н + + СО2.
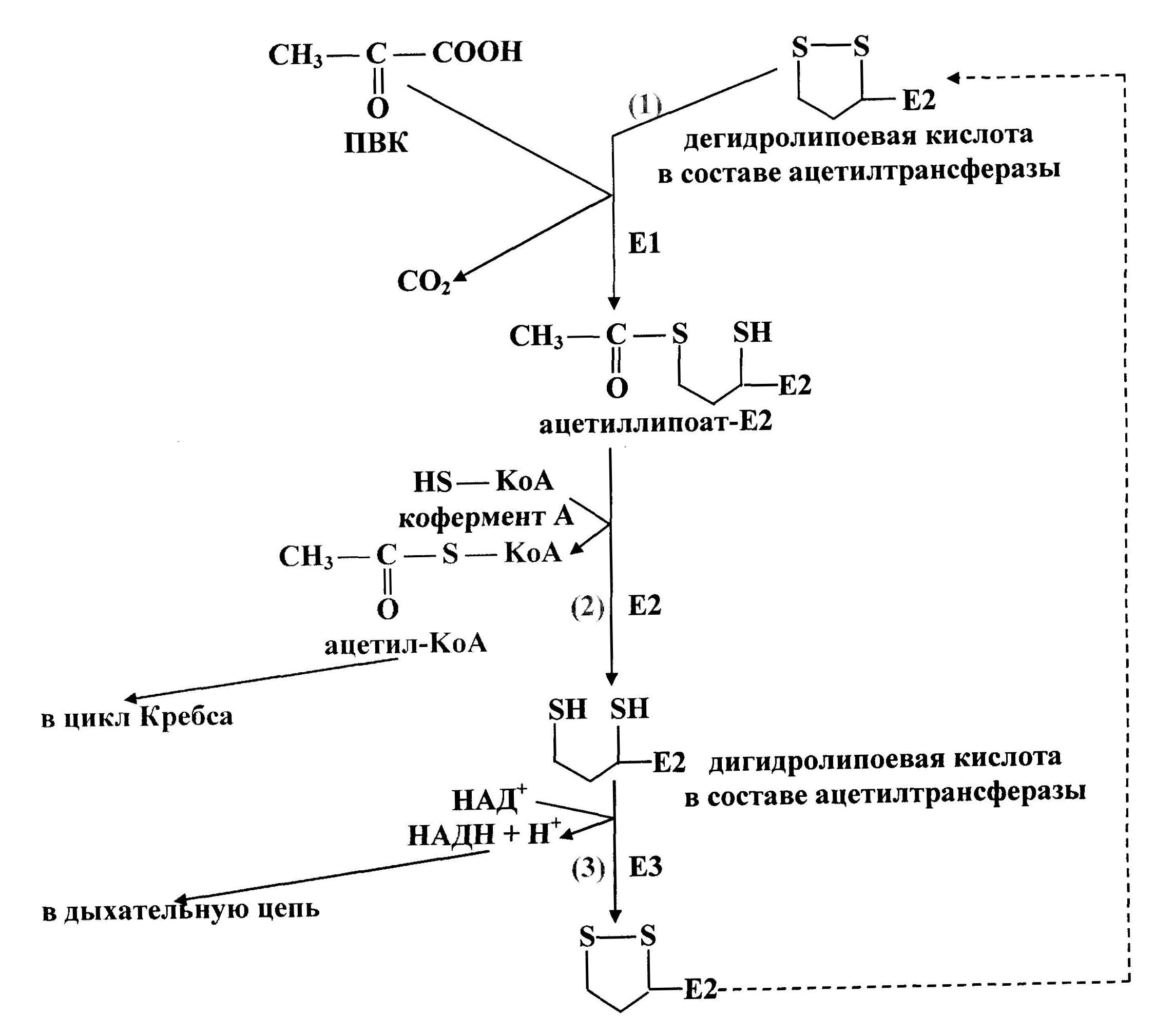
Рис. 37. Схема окислительного декарбоксилирования пирувата
По механизму «обратной связи» работу пируватдегидрогеназного комплекса ингибируют конечные продукты окислительного декарбоксилирования — ацетил-KоА, НАДН+Н + , а также АТФ. Увеличивают активность комплекса пировиноградная кислота и АДФ.
Также имеет место гормональная регуляция: инсулин увеличивает активность пируатдегидрогеназного комплекса, глюкагон — снижает.
Источник
Окислительное декарбоксилирование пирувата
Осуществляется при участии набора ферментов, объединенных в пируватдегидрогеназный комплекс (ПВДГК).Это мультиферментная система, которая включает 3 фермента и 5 коферментов (все они являются водорастворимыми витаминами) (рис. 6.4).
Е1— пируватдекарбоксилаза. Коферментом является активная форма витамина В1, тиамина — ТПФ (тиаминпирофосфат).
Е2— дигидролипоилацетилтрансфераза. Коферментом является витаминоподобноевещество — липоевая кислота (липоил), которая может временно превращаться в дигидролипоил, присоединив 2 атома водорода. Липоил может также переносить ацетильные остатки.
С этим ферментом также работает активная форма пантотеновой кислоты — КоА-SH, которая принимает ацетильный остаток от липоевой кислоты.
Е3— дигидролипоилдегидрогеназа. Коферментом является ФАД — активная форма витамина В2, рибофлавина. С работой этого фермента связан также кофермент НАД + — активная форма витамина РР, никотиновой кислоты.
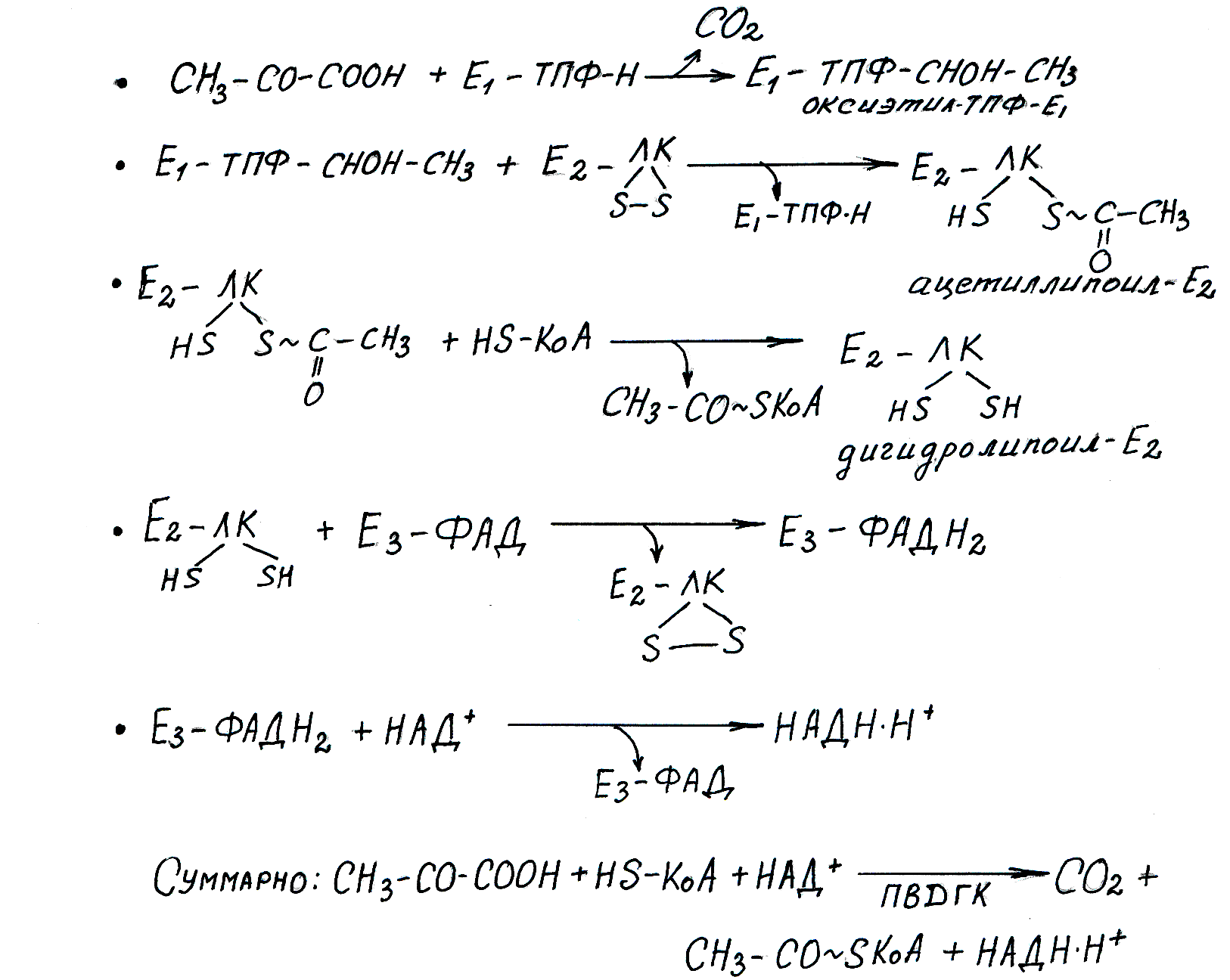
Рис. 6.4. Реакции окислительного декарбоксилирования пирувата
ПВГДК состоит из трех ферментов и пяти коферментов: Е1 – ТПФ-Н — пируватдекарбоксилаза (ТПФ — тиаминпирофосфат, активная форма витамина В1); Е 2 – ЛК — дигидролипоилацетилтрансфераза (ЛК — активная форма липоевой кислоты — витаминоподобного соединения); Е3 – ФАД — дигидролипоилдегидрогеназа (ФАД — флавинадениндинуклеотид — активная форма витамина В2); КоА-SH — активная форма пантотеновой кислоты; НАД + — никотинамидадениндинуклеотид — активная форма никотиновой кислоты. Схема регуляции ПВГДК предствалена на рисунке 6.5.
Таким образом, в результате образуются конечные продукты — СО2, атомы водорода для дыхательной цепи в составе НАДН·Н + и макроэргическое соединение ацетил-КоА. Лимитирующей реакцией в этом процессе является пируватдекарбоксилазная реакция. Поскольку фермент Е1в качестве кофермента использует ТПФ, при недостатке тиамина в пище нарушается окисление пирувата — процесса, который поставляет клеткам энергию. Возникает энергодефицит, что требует коррекции нарушения метаболизма с помощью тиамина.
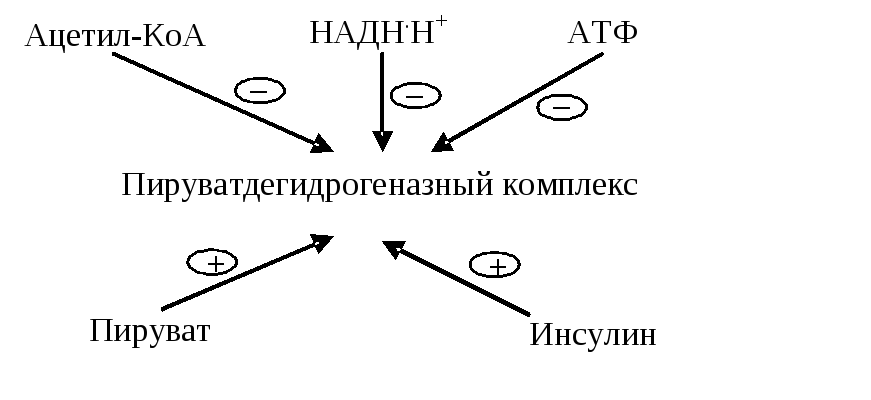
Рис. 6.5. Регуляция пируватдегидрогеназного комплекса
Лимоннокислый цикл Кребса, цикл трикарбоновых кислот (цтк)
Цикл лимонной кислоты локализован в матриксе митохондрий. Это циклический процесс из восьми последовательных реакций, в результате которых происходит декарбоксилирование и дегидрирование ацетил-КоА (универсального клеточного топлива) (рис. 6.6).
Цикл начинается с конденсации ацетил-КоА с 4-х углеродной кетокислотой — щавелевоуксусной (ЩУК). В результате образуется трикарбоновая кислота, цитрат. Изомеризация цитрата ведет к образованию изоцитрата. В ходе последовательных реакций изоцитрат декарбоксилируется и одновременно дегидрируется (фермент изоцитратДГ). Образовавшийся -кетоглутарат также декарбоксилируется и дегидрируется. Образовавшийся макроэрг сукцинил-КоА служит источником энергии для синтеза АТФ (субстратное фосфорилирование в цикле Кребса). В результате еще двух дегидрирований (ферменты сукцинатДГ и малатДГ) ЩУК регенерирует и запускает новый оборот цикла Кребса.
Таким образом, наряду с конечным продуктом обмена — СО2в четырех дегидрогеназных реакциях трижды восстанавливается НАД + (изоцитратДГ,-кетоглутаратДГ, малатДГ) и один раз восстанавливается ФАД (сукцинатДГ). Чтобы цикл мог функционировать, необходимо окислить эти коферменты, т. е. передать атомы водорода вдыхательную цепь,где происходит их окисление кислородом до воды.
Источник
43. Окислительное декарбоксилирование пировиноградной кислоты. Его биологическое значение.
Биологическая роль окислительного декарбоксилирования пирувата заключается в том, что оно является важным этапом катаболизма, позволяющим включаться в цикл Кребса тем веществам, при распаде которых образуется ПВК. Образовавшаяся молекула НАДН2 окисляется в длинной дыхательной цепи с образованием 3-х молекул АТФ. Окислительное декарбоксилирование пирувата протекает внутри митохондрий.
44. Какое участие кислорода в биологическом окислении? в каких случаях при тканевом дыхании возможно образование перекиси водорода н2о2? Каким образом организм ее обезвреживает?
Гонский (стр. 279-282)
45. Обмен пировиноградной кислоты в тканях.
Пировиноградная кислота (С3Н4O3) — α-кетопропионовая кислота. Используется обычно в виде солей — пируватов. Он является конечным продуктом метаболизма глюкозы в процессе гликолиза. Одна молекула глюкозы превращается при этом в две молекулы пировиноградной кислоты. Дальнейший метаболизм пировиноградной кислоты возможен двумя путями — аэробным и анаэробным. В условиях достаточного поступления кислорода, пировиноградная кислота превращается в ацетил-кофермент А, являющийся основным субстратом для серии реакций Пируват также может быть превращён в анаплеротической реакции в оксалоацетат. Оксалоацетат затем окисляется до углекислого газа и воды. Если кислорода недостаточно, пировиноградная кислота подвергается анаэробному расщеплению с образованием молочной кислоты При анаэробном дыхании в клетках пируват, полученный при гликолизе, преобразуется в лактат при помощи фермента
лактатдегидрогеназы и NADP в процессе лактатной ферментации, либо вацетальдегид и затем в этанол в процессе алкогольной ферментации. Пировиноградная кислота является «точкой пересечения» многих метаболических путей. Пируват может быть превращён обратно в глюкозу в процессе глюконеогенеза, или в жирные кислоты или энергию через ацетил-КоА,
в аминокислоту аланин, или в этанол. Например, работающая мышца выделяет в кровь наряду с молочной кислотой значительные количества аланина. Аланин образуется в мышце из пировиноградной кислоты путем трансаминирования. Из кровотока аланин поглощается печенью, превращается в пируват, а пируват используется для глюконеогенеза (глюкозо-аланиновый цикл, см. рис. 9.24).
Пировиноградная кислота содержится во всех тканях и органах и, являясь связующим звеном обмена углеводов, жиров и белков, играет важную роль в обмене веществ. Концентрация пировиноградной кислоты в тканях изменяется при болезнях печени, некоторых формах нефрита, раке, авитаминозах, особенно при недостатке витамина В1. Нарушение обмена пировиноградной кислоты приводит к ацетонурии.
Источник
Окислительное декарбоксилирование пирувата
Окисли́тельное декарбоксили́рование пирува́та — биохимический процесс, заключающийся в отщеплении одной молекулы углекислого газа (СО2) от молекулы пирувата и присоединения к декарбоксилированному пирувату кофермента А (КоА) с образованием ацетил-КоА; является промежуточным этапом между гликолизом и циклом трикарбоновых кислот. Декарбоксилирование пирувата осуществляет сложный пируватдегидрогеназный комплекс (ПДК), включающий в себя 3 фермента и 2 вспомогательных белка, а для его функционирования необходимы 5 кофакторов (КоА, НАД + , тиаминпирофосфат (ТПФ), ФАД и липоевая кислота (липоат)). Суммарное уравнение окислительного декарбоксилирования пирувата таково [1] :

У эукариот пируватдегидрогеназный комплекс локализован в митохондриях, у бактерий — в цитозоле. Образующийся в результате ацетил-КоА далее вовлекается в цикл Кребса [1] .
Окислительное декарбоксилирование пирувата — необратимый процесс. Образующийся в ходе этого процесса НАДН впоследствии отдаёт гидридный ион (Н — ) в дыхательную цепь, в которой при аэробном дыхании конечным акцептором электронов является кислород, а при анаэробном — другие окисленные соединения (например, сульфат, нитрат). Перенос электронов с НАДН на кислород даёт 2,5 молекулы АТФ на пару электронов. Необратимость реакции, осуществляемой пируватдегидрогеназным комплексом, была показана в исследованиях с применением радиоактивных изотопов: комплекс не может обратно присоединить меченый СО2 к ацетил-КоА с образованием пирувата [2] .
Помимо окислительного, существует неокислительное декарбоксилирование пирувата до ацетальдегида (и далее до этанола) и СО2. Этот процесс осуществляется ферментом пируватдекарбоксилазой, к нему способны многие растения, дрожжи и некоторые бактерии [3] .
Содержание
Коферменты
Некоторые коферменты ПДК



Комбинированное дегидрирование и декарбоксилирование пирувата до ацильной группы [en] , которая в дальнейшем войдёт в ацетил-КоА, осуществляется тремя различными ферментами, для функционирования которых необходимы 5 различных коферментов или простетических групп: тиаминпирофосфат (ТПФ), ФАД, кофермент А (КоА), НАД и липоат. Четыре из них являются производными витаминов: тиамина, или витамина В1 (ТПФ), рибофлавина, или витамина В2 (ФАД), ниацина, или витамина РР (НАД) и пантотеновой кислоты, или витамин В5 (КоА) [4] .
ФАД и НАД являются переносчиками электронов, а ТПФ известен также как кофермент пируватдекарбоксилазы, участвующей в брожении [4] .
Кофермент А имеет активную тиольную группу (—SH), которая имеет критическое значение для функционирования КоА в качестве переносчика ацильной группы в ряде метаболических реакций. Ацильные группы при этом ковалентно связываются с тиольной группой, образуя тиоэфиры. Из-за их относительно высокой стандартной свободной энергии гидролиза тиоэфиры обладают высокой способностью к переносу ацильных групп к различным молекулам-акцепторам. Поэтому ацетил-КоА иногда также называют «активированной уксусной кислотой» [4] [5] .
Пятый кофактор пируватдегидрогеназного комплекса, остаток липоевой кислоты — липоат, имеет две тиольные группы, которые могут подвергаться обратимому окислению с образованием дисульфидной связи (—S—S—), подобно тому, как это происходит между двумя остатками аминокислоты цистеина в белке. Из-за своей способности подвергаться окислению и восстановлению липоат может служить в качестве переносчика как электронов (или H + ), так и ацильных групп [4] .
Пируватдегидрогеназный комплекс
Пируватдегидрогеназный комплекс (ПДК)

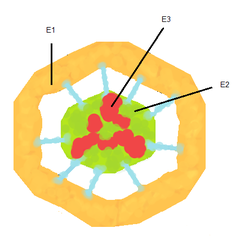
Пируватдегидрогеназный комплекс (ПДК) включает 3 фермента: пируватдегидрогеназу [en] (Е1), дигидролипоилтрансацетилазу (Е2) и дигидролипоилдегидрогеназу [en] (Е3). Каждый из этих ферментов присутствует в комплексе во множестве копий. Количество копий каждого фермента, а следовательно, и размер комплекса варьирует среди различных видов.
Комплекс ПДК млекопитающих достигает около 50 нм в диаметре, что более чем в 5 раз превышает диаметр целой рибосомы; эти комплексы достаточно велики, чтобы быть различимыми в электронный микроскоп. В ПДК коровы входят 60 идентичных копий Е2, которые формируют пентагональный додекаэдр ( коровая часть [en] комплекса) диаметром около 25 нм.
В кор ПДК у бактерии Escherichia coli входит 24 копии Е2. К Е2 присоединяется простетическая группа липоат (остаток альфа-липоевой кислоты) с аминокислотой лизином, которая связывается амидной связью с ε-аминогруппе остатка лизина, входящего в состав Е2. Е2 состоит из трёх функционально различных доменов: аминотерминального липоильного домена, содержащего остаток лизина, связывающийся с липоатом; центрального Е1— и Е3—связывающего домена; внутреннего корового ацилтрансферазного домена, содержащего активные центры ацилтрансферазы. У дрожжей в ПДК имеется единственный липоильный домен, у млекопитающих — два, а у E. coli — три. Домены Е2 связываются линкерными последовательностями, состоящими из 20—30 аминокислотных остатков, причём в них остатки аланина и пролина перемежаются с заряженными аминокислотыми остатками [6] .
С активным центром Е1 связывается ТПФ, а с активным центром Е3 — ФАД. Также в состав комплекса ПДК входят два регуляторных белка — протеинкиназа и фосфопротеинфосфатаза. Такая основная структура из Е1-Е2-Е3 оставалась консервативной в ходе эволюции. Комплексы такого устройства принимают участие и в других реакциях, например, окислении α-кетоглутарата в ходе цикла Кребса и окислении α-кетокислот, образующихся при катаболической утилизации разветвлённых аминокислот: валина, изолейцина, лейцина. У изученных видов Е3 ПДК идентичен Е3 двух вышеупомянутых комплексов. Примечательное сходство структур белков, кофакторов и механизмов реакций, осуществляемых этими комплексами, свидетельствует об общности их происхождения [1] . При прикреплении липоата к лизину Е2 образуется длинная, гибкая «рука», которая может перемещаться с активного центра Е1 в активные центры Е2 и Е3, то есть на расстояния предположительно 5 нм и более [7] .
Механизм
Окислительное декарбоксилирование пирувата включает несколько стадий:
- Стадия 1 идентична пируватдекарбоксилазной реакции. Первый атомуглерода (С-1) пирувата уходит в виде СО2, а С-2, в пирувате находящийся в альдегидной форме, прикрепляется к ТПФ в виде гидроксиэтильной группы (—СНОН—СН3). Первая стадия является наиболее медленной и поэтому ограничивает скорость всего процесса. Кроме того, на этом этапе комплекс ПДК проявляет свою субстратную специфичность. Эта реакция осуществляется пируватдегидрогеназой (Е1).
- Стадия 2. Гидроксиэтильная группа окисляется до карбоновой кислоты (ацетата). Два электрона, освобождаемых при этой реакции, идут на восстановление связи —S—S— липоильной группы Е2 до двух тиольных (—SH) групп.
- Стадия 3. Ацетильный остаток, образующийся в ходе окислительно-восстановительной реакции на стадии 2, сначала связывается тиоэфирной связью с липоильной —SH-группой, а затем переносится на КоА с образованием ацетил-КоА. Таким образом, энергия окисления идёт на образование высокоэнергетического тиоэфира ацетата. Стадии 2 и 3 катализируются дигидролипоилтрансацетилазой (Е2).
- Стадия 4 и стадия 5 катализируются дигидролипоилдегидрогеназой (Е3). В ходе этих двух последних реакций восстановленный липоиллизин снова возвращается в окисленную форму, который в дальнейшем может участвовать в следующем цикле окислительного декарбоксилирования пирувата. Электроны, изначально принадлежавшие гидроксиэтильной группе, при этом переносятся с липоиллизина сначала на ФАД с образованием ФАДH2, а потом на НАД + с образованием НАДН + H + [8] .

Центральную роль в реакции, осуществляемой комплексом ПДК, играют липоиллизиновые «руки» Е2, способные «раскачиваться» и забирать два электрона от Е1, а также ацетильную группу, образовавшуюся из пирувата, и доставлять электроны к Е3. Все эти ферменты и коферменты собраны в комплекс, благодаря чему промежуточные соединения могут вступать в необходимые реакции быстро и не диффундируя с поверхности ферментного комплекса. За счёт этого промежуточные соединения не покидают комплекса, и поддерживается очень высокая локальная концентрация субстрата Е2. Это также предотвращает перехватывание активированной ацетильной группы другими ферментами, использующими её в качестве субстрата [8] .
Органические соединения, содержащие мышьяк, являются ингибиторами ПДК, поскольку взаимодействуют с восстановленными в ходе окислительного декарбоксилирования пирувата тиольными группами липоильной группы Е2 и блокируют их нормальную работу [9] .
Регуляция
У млекопитающих ПДК сильно подавляется АТФ, а также продуктами реакции: ацетил-КоА и НАДН. Аллостерическое подавление окисления пирувата значительно усиливается в присутствии длинноцепочечных жирных кислот. АМФ, КоА и НАД + , накапливающиеся тогда, когда в цикл Кребса поступает слишком мало ацетата, аллостерически активируют комплекс ПДК. Таким образом, ферментный комплекс подавляется, когда имеется достаточно ацетил-КоА или сырья (жирные кислоты) для осуществления альтернативных путей образования ацетил-КоА, а отношения [АТФ]/[АДФ] и [НАДН]/[НАД + ] достаточно велики. Напротив, при большой потребности в энергии и необходимости большего количества ацетил-КоА для функционирования цикла Кребса ПДК активируется [10] .
У млекопитающих к этим аллостерическим механизмам добавляется второй уровень регуляции: ковалентная модификация белка. Комплекс ПДК подавляется обратимым фосфорилированием по специфическим остаткам серина на одной из двух субъединиц E1. Ранее отмечалось, что, помимо субъединиц E1, E2 и E3 у млекопитающих в комплекс ПДК входят два регуляторных белка, единственным назначением которых является регуляция активности комплекса. Специфичная протеинкиназа фосфорилирует и тем самым инактивирует E1, а специфичная фосфопротеинфосфатаза удаляет фосфатные группы путём гидролиза и тем самым активирует E1. Киназа аллостерически активируется АТФ: когда концентрация АТФ велика (что свидетельствует о достаточном количестве энергии в клетке), комплекс ПДК инактивируется фосфорилированием E1. Когда [АТФ] понижена, активность киназы снижается, и фосфатаза убирает фосфатные группы с E1, активируя комплекс [11] .
Комплекс ПДК растений, располагающийся в матриксе митохондрий и пластидах, подавляется продуктами его активности — НАДН и ацетил-КоА. Растительный митохондриальный фермент также регулируется обратимым фосфорилированием: пируват подавляет киназу, активируя ПДК, а NH4 + стимулирует киназу и инактивирует комплекс. У E. coli ПДК регулируется аллостерически по схожему с млекопитающими механизму, однако, по-видимому, не регулируется фосфорилированием [11] .
Клиническое значение
Четыре витамина (тиамин, рибофлавин, ниацин, пантотеновая кислота), из которых образуются коферменты ПДК, обязательно должны присутствовать в рационе человека [4] . Кроме того, мутации генов, кодирующих субъединицы ПДК, а также недостаток тиамина в пище могут иметь очень серьёзные последствия. Животные, испытывающие недостаток тиамина, не могут нормально окислять пируват. Особенно это важно для мозга, который обычно получает энергию при аэробном окислении глюкозы, а этот процесс обязательно включает окисление пирувата.
Бери-бери — заболевание, развивающееся при недостатке тиамина — характеризуется расстройством функций нервной системы. Эта болезнь обычно встречается в популяциях людей, чей рацион состоит в основном из белого (очищенного) риса, лишённого шелухи, в которой содержится большая часть тиамина риса. Недостаточность тиамина может также развиться у людей, постоянно употребляющих алкоголь, так как большая часть получаемой ими энергии приходится на «пустые калории» очищенного спирта, лишённого витаминов. Повышенное содержание пирувата в крови часто является индикатором нарушений в окислении пирувата из-за одной из вышеперечисленных причин [12] .
Другие пути преобразования пирувата
У некоторых микроорганизмов преобразование пирувата в ацетил-КоА (или другие продукты) может осуществляться и другими способами, помимо вышеописанного (комплекс ПДК используется аэробами). Такими преобразованиями могут быть:
- пируват + КоА + ФАД → ацетил-КоА + ФАДH2 + СО2. Реакция катализируется пируват:ферредоксин оксиредуктазой[en], характерна для анаэробов, в частности, клостридий. Из ФАДH2 далее при участии гидрогеназы[en] образуется молекулярный водород.
- пируват + КоА → ацетил-КоА + формиат. Фермент — пируватформиатлиаза[en], реакция характерна для Enterobacteriaceae, фотобактерий[en] и некоторых фототрофов.
- пируват → ацетальдегид + СО2. Фермент — пируватдекарбоксилаза, реакция характерна для дрожжей [13] .
Источник