Метаболизм витамина В12

Весь контент iLive проверяется медицинскими экспертами, чтобы обеспечить максимально возможную точность и соответствие фактам.
У нас есть строгие правила по выбору источников информации и мы ссылаемся только на авторитетные сайты, академические исследовательские институты и, по возможности, доказанные медицинские исследования. Обратите внимание, что цифры в скобках ([1], [2] и т. д.) являются интерактивными ссылками на такие исследования.
Если вы считаете, что какой-либо из наших материалов является неточным, устаревшим или иным образом сомнительным, выберите его и нажмите Ctrl + Enter.
Витамин В12 (кобаламин) — входит в состав продуктов только животного происхождения: мясо, печень, молоко, яйца, сыр и другие (витамин в тканях животных является производным бактерий). Под влиянием кулинарной обработки и протеолитических ферментов желудка он высвобождается и быстро связывается с «R-binders» (транс-кобаламины I и III) — белками с быстрой (rapid) электро-форетической подвижностью, по сравнению с внутренним фактором; в меньшей степени витамин В12 соединяется с внутренним фактором (ВФ, фактор Касла) — гликопротеином, вырабатываемым париетальными клетками фундальной части и тела желудка. Под влиянием протеаз панкреатического сока комплекс R-витамин В,2 разрушается и высвобождается витамин В12, который соединяется с внутренним фактором, образуя комплекс витамин В12-ВФ. Последний днмеризуется и в подвздошной кишке связывается со специфическими рецепторами. В присутствии ионов кальция и при рН 7,0 этот комплекс расщепляется и витамин В12 проникает в митохондрии клеток слизистой оболочки кишки. Отсюда витамин В12 проникает в кровь, где соединяется с транспортным белком транскобаламином II (ТК II), который доставляет витамин к тканям-мишеням — гепатоцитам, гемопоэтическим клеткам и другим.
Высвобождение витамина В12 из комплекса ТКП-В12 в клетке происходит в 3 этапа:
- связывание комплекса с рецепторами клетки;
- его эндоцитоз;
- лизосомальный гидролиз с высвобождением витамина.
Часть витамина В12 в сыворотке крови связывается с кобалофилинами («R-binders») — транскобаламинами I и III. Эти В12-связывающие гликопротеины высвобождают его только в печени. При наличии большого количества витамина В12 около 1 % его может проникать в кровь путем пассивной диффузии.
Основным депо витамина В12 является печень, в 1 г которой содержится 1 мкг витамина В12. У здоровых доношенных детей резервы витамина В12 в печени составляют 20-25 мкг, депо резко истощается к году. Суточная потребность в витамине В12 у ребенка грудного возраста составляет 0,1 мкг, а у взрослого — 5-7 мкг. В 100 мл женского молока содержится 0,11 мкг витамина В12.
Витамин В12 в основном выделяется с желчью, его потеря происходит также с калом; в сутки теряется 0,1 % от всего депонированного витамина. Доказано существование кишечно-печеночного кругооборота витамина В12— около 3/4 выделенного с желчью витамина вновь реабсорбируется. Этим объясняется развитие мегалобластной анемии через 1-3 года после полного прекращения поступления витамина В12 в организм. Физиологические потери витамина с мочой крайне незначительны.
Источник
Метаболизм витамина в12 и фолатов и их роль в кроветворении
Кобальтсодержащий витамин В12 (кобаламин)содержится в продуктах питания животного происхождения, богатых белком – мясе, яйцах, сыре, молоке, особенно много его в печени и почках.
После освобождения от пищевого белка витамин В12связывается сR-белком слюны, а затем соединяется свнутренним фактором Касла (гастромукопротеином)– белком, секретируемым париетальными клетками слизистой оболочки желудка. Образовавшийся комплекс абсорбируется через специфические рецепторы подвздошной кишки, количество которых лимитирует скорость поступления кобаламина в организм. В кровотоке витамин В12переносится специализированными транспортными белкамитранскобаламинами. Конечными этапами переноса кобаламина являются гемсинтезирующие клетки костного мозга и печени.
Главным депо витамина В12является печень, в 1 г которой содержится 1 мкг кобаламина. Суточная потребность в нем для взрослого человека составляет 5-7 мкг. Выделяется из организма витамин В12с желчью, его потеря происходит с калом. За сутки теряется 0,1% от всего количества депонированного витамина. Доказано существование кишечно-печеночной циркуляции кобаламина, при этом около 75% выделяемого с желчью витамина вновь реабсорбируется, чем объясняется развитие В12ДА через 1-3 года после полного прекращения поступления его в организм.
Фолатышироко распространены в пищевых продуктах животного и растительного происхождения, они содержатся в печени, дрожжах, мясе, шпинате, шоколаде, сырых овощах и фруктах. Более 50% фолиевой кислоты может разрушаться в процессе тепловой кулинарной обработке пищи, чем объясняется ее дефицит у лиц, потребляющих вареные продукты. Запасфолиевой кислотыу человека составляет 5-20 мг. Суточная потребность в ней – 50-100 мкг.
Фолиевая кислота всасывается в двенадцатиперстной кишке и проксимальном отделе тощей кишки. Способность кишечника всасывать фолаты превышает суточную потребность в них. В плазме крови они связываются с различными белками (2-макроглобулином, альбумином) и транспортируется в печень, где депонируются в виде полиглутаматов, небольшое количество экскретируется с мочой. Запасы фолиевой кислоты истощаются уже через 3-4 месяца после прекращения поступления фолатов в организм.
Этиология и патогенез мегалобластных анемий
Наиболее частыми причинами развития дефицита витамина В12являются:
отсутствие внутреннего фактора Касла (аутоиммунный атрофический гастрит типа А, гастрэктомия);
поражение тонкого кишечника (энтерит, резекция тонкой кишки, тропическая спру, целиакия, полипоз, злокачественные новообразования, синдром Имерслунд-Гресбека).
Недостаточное поступление с пищей:
строгая вегетарианская диета.
инвазия широким лентецом (дифиллоботриоз);
патологическая микрофлора кишечника при наличии дивертикулеза либо «слепой петли».
Наследственный дефицит транскобаламина.
Основной причиной развития дефицита витамина В12является хронический атрофический гастрит (тип А), при котором прекращается либо уменьшается синтез внутреннего фактора в результате аутоиммунного поражения железистых структур слизистой оболочки желудка. Иммунные механизмы могут играть ведущую роль в развитии В12ДА у лиц молодого возраста. Нередко она сочетается с другими аутоиммунными заболеваниями (зоб Хашимото, гемолитическая анемия).
Существенное значение для развития дефицита витамина В12имеетнаследственная предрасположенность, 20-30% больных имеют родственников с В12ДА.
Причиной развития В12ДА может бытьконкурентное потребление кобаламина в кишечнике патогенной микрофлорой и паразитами, использующими его для синтеза ДНК бактерий.
Причинами развития дефицита фолиевой кислотыв организме могут быть:
1. Снижение содержания в пище:
алкоголизм, голодание, «бутербродный» рацион питания;
длительная кулинарная обработка пищи.
2. Нарушение всасывания:
хронический энтерит, резекция тонкой кишки, диабетическая энтеропатия, тропическая и нетропическая спру;
недоношенные дети, находящиеся на искусственном вскармливании.
3. Повышение потребности:
гемолитические анемии, лейкозы, злокачественные новообразования, туберкулез, эксфолиативный дерматит, гипертиреоз.
4. Уменьшение запасов в печени:
алкоголизм, цирроз печени, гепатоцеллюлярная карцинома.
5. Прием лекарственных препаратов:
Процессы всасывания в пищеварительном тракте и поступления фолатов в организм часто нарушены при алкоголизме, поскольку в этом случае, как правило, имеет место недостаточное поступление продуктов растительного и животного происхождения. Кроме того, алкоголь нарушает метаболизм фолатов. Аналогичная ситуация развиваетсяу малообеспеченных категорий населения, особенно, у престарелых лиц.
Повышение потребности в фолатах формируется в период интенсивного ростав детском и юношеском возрасте, а также при беременности.
ФДА могут формироваться при приеме лекарственных препаратов, разными механизмами воздействующих на метаболизм фолатов и синтез ДНК. К их числу относятся ингибиторы синтеза ДНК прямого действия (азатиоприн, 6-меркаптопурин, 5-фторурацил, цитозинарабинозид, гидроксикарбамид, прокарбазин), антагонисты фолиевой кислоты (метотрексат, триметоприм, триамтерен), закись азота, противосудорожные средства (фенитоин, примидон, фенобарбитал).
Патогенез МБАобусловлен дефицитом витамина В12и фолатов в организме, когда нарушаются метаболические процессы, в которых они принимают участие (Рис. 2).
Витамин В12в организме участвует в процессесинтеза нуклеиновых кислот. При дефиците его замедляется синтез ДНК во всех ядросодержащих клетках, в первую очередь, гемопоэтических, которые обладают наиболее высокой пролиферативной активностью, что ведет к формированию МБА. Кроме того, дефицит кобаламина сопровождается нарушениемсинтеза и распада жирных кислот в нервной тканис накоплением токсичных для нервной системы пропионовой и метилмалоновой кислот, приводящих к поражению задних и боковых столбов спинного мозга, а также к нарушению образования миелина с повреждением аксонов.
Фолаты, как и витамин В12, занимают ключевое положение во многих видах клеточного метаболизма, включаясинтез аминокислот, нуклеиновых кислот и пуриновых оснований, необходимых для образования ДНК в активно пролиферирующих клетках.
Метаболические процессы с участием кобаламина и фолатов тесно связаны между собой. Проникновение фолатов в клетку является витамин В12-зависимым процессом. При дефиците кобаламина расстраивается и метаболизм фолиевой кислоты с нарушением синтеза ДНК и появлением мегалобластического кроветворения, при этом
Источник
Обмен витамина в12 биохимия
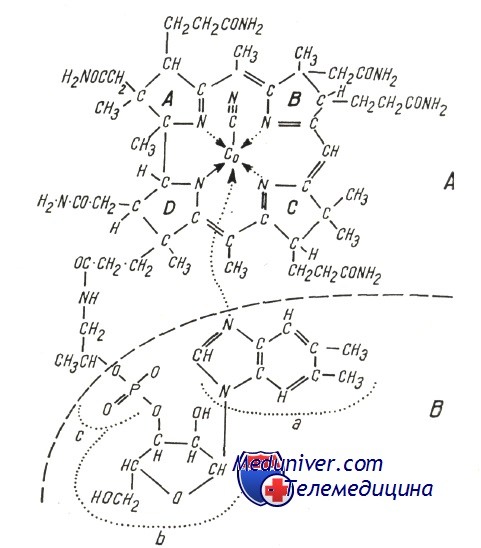
Молекула витамина В12 (цианкобаламин) состоит из двух частей:
а) хориновой — порфирин, в котором 4 восстановленных пирола связаны тремя мостиками метена и одной прямой связью, при этом к внешним позициям прикреплены радикалы, такие как аминоэтанол, аминопропанол и метил, в то время как в центре расположен атом кобальта, с которым связана группировка циан (свреху) и нуклеотидная часть (под хориновым планом);
б) нуклеотидной, в состав которой входят 5,6-диметилбензимидазол (а), рибоз (b) и фосфат (с) (которая осуществляет связь с пропионовым радикалом пирола D). Заменой связанных с кобальтом радикалов образуются различные производные (естественные и синтетические); при этом коферментной активностью обладают лишь высокозаменяемые произволные, в которых группа —С—N заменяется радикалом 5′-дезоксиаденозил или метил.
В качестве кофермента 5′-дезоксиадеознилкобаламин является межмолекулярным (реакция восстановления) или внутримолекулярным (реакция изомеризации) переносчиком водорода. Был обнаружен целый ряд реакций подобного типа, однако из них, у млекопитающих и человека, доказано наличие лишь реакции изомеризации метил-малонил-СоА в сукцинил-СоА в процессе метаболизма пропионовой кислоты (Silber и Moldow).
Реакцию восстановления рибонуклеотидов в дезоксирибонкулеотиды, обнаруженную в отдельных микроорганизмах, посредством которой можно было бы объяснить мегалобластоз за счет дефицита BJ2 независимым от метаболизма фолатов путем, еще не удалось доказать у человека (тем не менее добавление тимидина может преобразовать мегалобласты в нормобласты только при недостатке фолиевой кислоты; исключаются случаи дефицита витамина В12).
В бактериях метилкобаламин участвует в различных реакциях синтезирования, выполняя роль переносчика радикала метил. У человека доказано наличие лишь реакции метилирования гомоцистеина (в одном из путей синтезирования метионина). Поскольку источником группы метил является метилтетрагидрофолат, эта реакция составляет одновременно путь к «регенерации» тетрагидрофолиевой кислоты (FH4) из метил-тетрагидрофолиевой кислоты, что служит признанным в настоящее время объяснением мегалобластоза за счет дефицита витамина В12.
При этом теория «капкан метилфолата» (Nixon и Bertino) составляет другое последствие дефицита витамина В12, заключающееся в недостаточной задержке клетками метилтетрагидрофолиевой кислоты (Тисман и Герберт).
Дозировка витамина В12 осуществляется бактериологическими методами, при этом применяется либо фотометрическое измерение роста численного показателя отдельных микроорганизмов, находящихся в зависимости от этого витамина (Lactobacillus leishmanii, Euglena gracilis и пр.), когда вступают в контакт с материалом реакции, либо метод радиоактивных изотопов.
Кругооборот витамина В12 в организме человека
Витамин В12 синтезируется отдельными микроорганизмами, находящимися в почве, воде, на кормовых растениях или в пищеварительном тракте животных, далее витамин поступает в ткани (в частности в железы, печень, мышцы, также, яйца, в меньшей мере в молоко), которые составляют основной источник для человека.
Из 5—30 мкг среднесуточного подвоза с пищей поглощаются лишь 1—5 мкг, остальное количество выделяется с испражнениями. За сутки теряются примерно 2,5 мкг витамина В12 (Hellmuth), составляющие, следовательно, суточную потребность. Его передача плоду, в период беременности, различные гиперметаболические состояния, в период роста и пр. увеличивают потребность.
Поглощение витамина В12 обусловлено наличием внутреннего фактора (ВФ), который связывает находящийся в пищевых продуктах витамин В12 и затем переносит его к кишечным рецепторам.
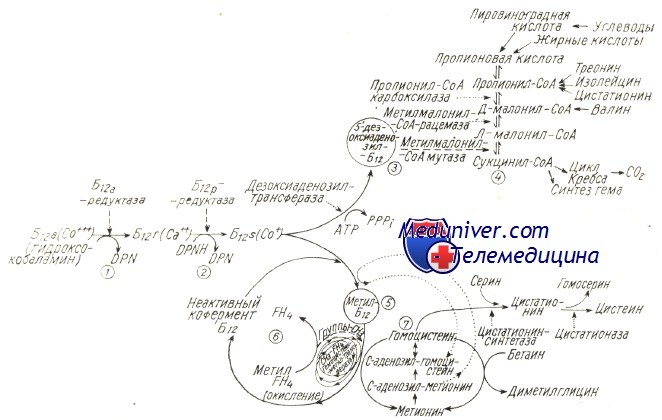 Схематическое изображение синтеза коферментов и их метаболической активности.
Схематическое изображение синтеза коферментов и их метаболической активности.
Приводятся два последовательных восстановительных процесса гидроксокобаламина (1), (2), образование 5′-дезоксиаденозил-В12 (3) и его участие в метаболизме пропионовой кислоты (4); образование метип-В12 (5), его «каталитическая» функция в переносе группы — СН3 от метил-FH4 к гомоцистеину (пусковая роль s-аденозилметионина в реакции и «регенерации’» кофермента), также взаимосвязь с метаболизмом фолатов (6) и серных аминокислот (7).
Внутренний фактор это, по существу, гликопротеид, который, у человека, выделяется пристеночными клетками слизистой оболочки дна желудка. Этот фактор отличается свойством осуществлять специфическую связь (при кислом рН) того небольшого количества В12, которое выделяется в результате пептического пищеварения.
Далее, при нейтральном рН и наличии Са2+ комплекс ВФ-В12 закрепляется на специальных рецепторах слизистой оболочки подвздошной кишки, где витамин диссоциируется а затем, с помощью еще не выведенного механизма, переходит в воротновенную кровь (Састле).
При введении большого количества витамина В12 его поглощение осуществляется путем пассивной диффузии (с быстрым поступлением в кровь) в размере примерно 1% назначенной дозы (Hellmuth).
Перенос с плазмой витамина В12 осуществляется посредством транскобаламина (прежде называемого транскобаламин II), связывающего поглощаемый из кишечника и частично эндогенный витамин В12, равно как кобалофилина (современное название «белков вида Р », включающих транскобаламин I и III), переносящего наибольшую часть эндогенного витамина В12. При хронической гранулоцитной лейкемии, а иногда и гепатомах показатель кобалофилина значительно растет. Недавно был выявлен внутриклеточный белок, связывающий кобаламин (Stenman).
Нормальная концентрация витамина В12 в плазме колеблется от 200 до 900 пг/мл.
Запасы витамина В12 в организме взрослого человека составляют примерно 2000—5000 мкг, причем в основном он сохраняется в печени (примерно 1000 мкг) и в меньшей мере в почках; мышцы содержат примерно 3000 мкг этого витамина.
Витамин В12 выделяется с мочой в незначительном количестве (примерно (0,25 мкг/сутки). Видимо его выделение через кишечник составляет основную причину потерь (доказано наличие кишечно-печеночного кругооборота с повторным поглощением примерно 3/4 выделяемого с желчью витамина В12), к этому следует добавить выпадение клеток желудочно-кишечного эпителия, выделения пищеварительной системы и пр.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Источник