Цикл Кребса — схема реакций, ферменты, коферменты, энергетический баланс одного оборота. Тканевые особенности в детском возрасте, Регуляция.
Цикл Кребса.
ЩУКа «съела» ацетат,
Будет он — изоцитрат.
Водороды отдав НАД,
Этому безмерно рад
Альфа -кето- глутарат.
НАД похитит водород,
С коэнзимом А спешат,
В сукциниле появилась,
Сразу АТФ родилась.
И остался сукцинат.
Вот добрался он до ФАДа —
Водороды тому надо.
Стал он просто фумарат.
Фумарат воды напился,
Да в малат и превратился.
Тут к малату НАД пришел,
ЩУКа снова объявилась
И тихонько затаилась
Ферменты в этой схеме есть.
Коферменты — это НАД, НАДФ, АТФ, ГТФ? Тогда есть.
Схема:
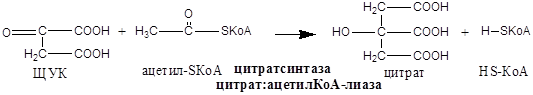
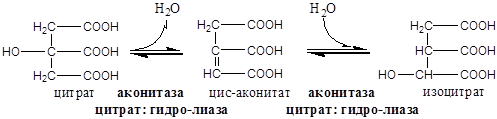
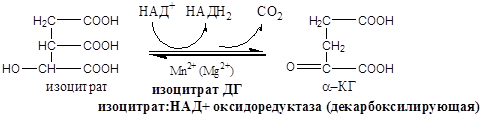
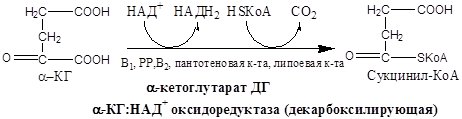
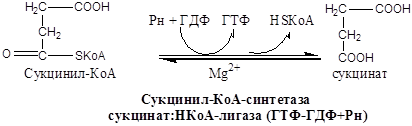
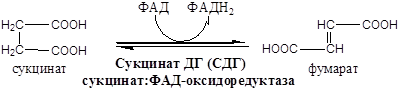
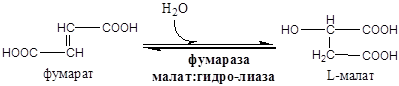
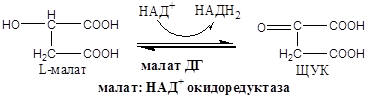
Образовавшиеся молекулы ЩУК реагируют с новой молекулой Ацетил-КоА и цикл повторяется вновь.
Энергетический баланс одного оборота:3 НАДН2 + 1 ФАДН2 (направляются далее в дыхательную цепь окислительного фосфорилирования) + 1 ГТФ (НАДН2 -> 3 АТФ, ФАДН2 -> 2 АТФ, ГТФ -> 1 АТФ) = 12 АТФ.
Регуляция ЦТК: 4 регуляторных фермента: цитратсинтазы, изоцитрат ДГ, α-КГ ДГ и СДГ. ЦТК ингибируется в основном НАДН2 и АТФ, которые являются продуктами ЦТК и цепи окислительного фосфорилирования. Активируют ЦТК в основном НАД + и АДФ.
Оксидазный путь использования кислорода в клетке — митохондриальное окислительное фосфорилирование. Состав дыхательных комплексов редокс-цепи, локализация и функции, тканевые особенности в детском возрасте. Регуляция.
Оксидазный путь использования кислорода в клетке:
Протекает в митохондриях, потребляет 90% О2 и обеспечивает процесс окислительного фосфорилирования.
Окислительное фосфорилирование — синтез АТФ из АДФ и Н3РО4 за счет энергии движении электронов по дыхательной цепи.
Оно является основным источником АТФ в аэробных клетках
Окислительное фосфорилирование состоит из процессов окисления и фосфорилирования.
1) Процесс окисления
Процесс окисления происходит при движении электронов по дыхательной цепи от субстратов тканевого дыхания на кислород. Дыхательная цепь окислительного фосфорилирования состоит из 4 белковых комплексов, встроенных во внутреннюю мембрану митохондрий и небольших подвижных молекул убихинона и цитохрома С, которые циркулируют в липидном слое мембраны между белковыми комплексами.
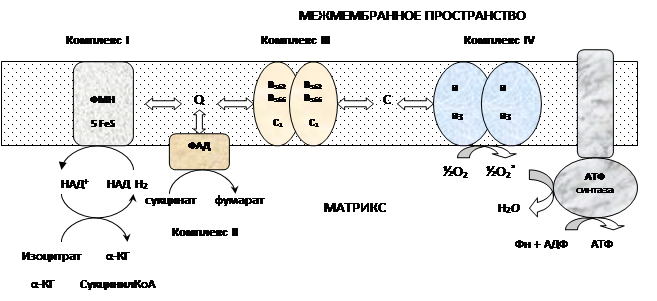
a. Комплекс I – НАДН2 дегидрогеназный комплекс–самый большой из дыхательных ферментных комплексов, в качестве коферментов содержит ФМН и 5 железосерных (Fe2S2 и Fe4S4) белков.
b. Комплекс II – СДГ. В качестве коферментов содержит ФАД и железосерный белок.
c. Комплекс III – Комплекс b-c1 (фермент QH2 ДГ). Каждый мономер содержит 3 гема, связанных с цитохромами b562, b566, с1, и железосерный белок.
d. Комплекс IV – Цитохромоксидазный комплекс. Каждый мономер содержит 2 цитохрома (а и а3) и 2 атома меди.
e. Коэнзим Q (убихинон). Переносит по 2Н + и 2е — .
f. Цитохром С. Периферический водорастворимый мембранный белок. Содержит молекулу гема.
Этапы движения е — по дыхательной цепи
a. 2е — от НАДН2, проходят через I комплекс (ФМН→SFe белок) на КоQ, высвобождаемая при этом энергия обеспечивает перекачку Н + .
b. КоQ с 2е — забирает у воды 2Н + из матрикса и превращается в КоQН2 (восстановление КоQ проходит также с участием комплекса II).
c. КоQН2 переносит 2е — на комплекс III, а 2Н + в межмембранное пространство.
d. Цитохром С переносит е — c III комплекса на IV комплекс.
e. IV комплекс сбрасывает е — на О2, высвобождаемая при этом энергия обеспечивает перекачку Н + .
Образовавшийся на внутренней мембране митохондрий электрохимический потенциал используется для:
a. фосфорилирования АДФ в АТФ;
b. транспорта веществ через мембрану митохондрий;
2) Процесс фосфорилирования
Процесс фосфорилирования осуществляется АТФ-синтетазой (Н + -АТФ-аза), которая потребляет 40-45% свободной энергии, выделившейся при окислении. Н + -АТФ-аза интегральный белок внутренней мембраны митохондрий, она состоит из 2 белковых комплексов F и F1.
a. Гидрофобный комплекс F погружён в мембрану и служит основанием, которое фиксирует АТФ-синтазу в мембране. Он состоит из нескольких субъединиц, образующих канал, по которому протоны переносятся в матрикс.
b. Комплекс F1 выступает в митохондриальный матрикс. Он состоит из 9 субъединиц (3α, 3β, γ, δ, ε). Субъединицы α и β уложены попарно, образуя «головку»; между а- и β-субъединицами располагаются 3 активных центра, в которых происходит синтез АТФ; γ, δ, ε – субъединицы связывают комплекс F1, с F.
АТФ-синтетаза обеспечивает обратимое взаимопревращение энергии электрохимического потенциала и энергии химических связей.
Электрохимический потенциал внутренней мембраны заставляет Н + двигаться из межмебранного пространства по каналу АТФ-синтазы в матрикс митохондрий. При каждом переносе протонов через канал Fo энергия электрохимического потенциала расходуется на поворот стержня, в результате которого циклически изменяется конформация а- и β-субъединиц и все 3 активных центра, образованных парам α- и β-субъединиц, катализируют очередную фазу цикла: 1) связывание АДФ и Н3РО4; 2) образование фосфоангидридной связи АТФ; 3) освобождение конечного продукта АТФ.
Дата добавления: 2016-06-29 ; просмотров: 7751 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Источник
44. Цикл Кребса, биологическая роль, основные реакции. Ферменты цикла Кребса.
цикл Кребса – представляет собой замкнутую систему биохимических окислительно-восстановительных реакций. Цикл назван по имени английского биохимика Ханса Кребса, который постулировал и экспериментально подтвердил основные реакции аэробного окисления. За проведенные исследования Кребс получил Нобелевскую премию (1953). Цикл имеет еще два названия:
цикл трикарбоновых кислот, так как он включает реакции превращения трикарбоновых кислот (кислот, содержащих три карбоксильные группы);
цикл лимонной кислоты, так как первой реакцией цикла является образование лимонной кислоты.
Цикл Кребса включает 10 реакций, четыре из которых окислительно-восстановительные. В ходе реакций освобождается 70% энергии.
Чрезвычайно велика биологическая роль этого цикла, поскольку это общий конечный пункт окислительного распада всех основных пищевых продуктов. Это главный механизм окисления в клетке, образно его называют метаболическим «котлом». В процессе окисления топливных молекул (углеводов, аминокислот, жирных кислот происходит обеспечение организма энергией в виде АТФ. Топливные молекулы вступают в цикл Кребса после превращения в ацетил-Ко-А.
Кроме того, цикл трикарбоновых кислот поставляет промежуточные продукты для процессов биосинтеза. Этот цикл происходит в матриксе митохондрий. Рассмотрим реакции цикла Кребса
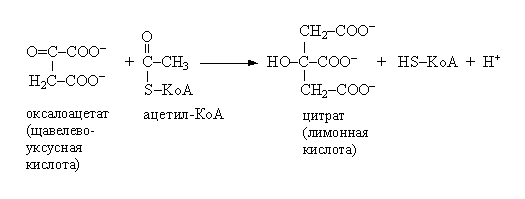
Цикл начинается с конденсации четырехуглеродного компонента оксалоацетата и двухуглеродного компонента ацетил-Ко-А. Реакция катализируется цитратсинтазой и представляет собой альдольную конденсацию с последующим гидролизом. Промежуточным продуктом является цитрил-Ко-А, который гидролизуется на цитрат и КоА:
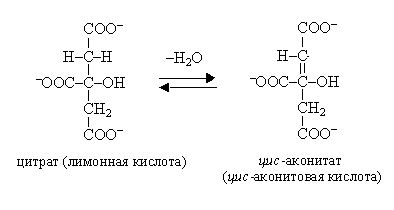
Данный процесс является реакцией дегидратации, катализируется ферментом аконитазой.

I 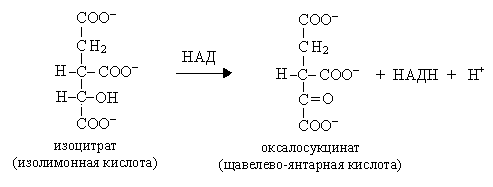 II. Данный процесс является реакцией гидратации, катализируется ферментом аконитазой.
II. Данный процесс является реакцией гидратации, катализируется ферментом аконитазой.
I 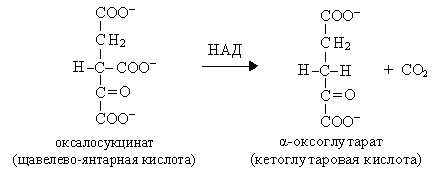 V. Это первая окислительно-восстановительная реакция.
V. Это первая окислительно-восстановительная реакция.
Реакции 4 и 5 представляют собой окислительное декарбоксилирование, катализируются изоцитратдегидрогеназой, промежуточным продуктом реакций является оксалосукцинат.
V 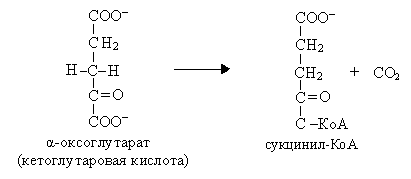 I. Эта реакция также является реакцией окислительного декарбоксилирования, т.е. это вторая окислительно-восстановительная реакция:
I. Эта реакция также является реакцией окислительного декарбоксилирования, т.е. это вторая окислительно-восстановительная реакция:
α-Оксоглутарат + НАД + КоА Сукцинил-КоА + СО2 + НАДН
Реакция катализируется α-оксоглутаратдегидрогеназным комплексом, состоящим из трех ферментов:
В 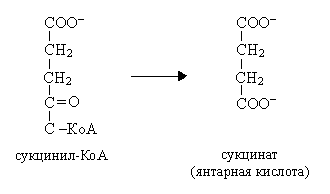 сукциниле имеется связь, богатая энергией. Расщепление тиоэфирной связи сукцинил-КоА сопряжено с фосфорилированием гуанозиндифосфата (ГДФ):
сукциниле имеется связь, богатая энергией. Расщепление тиоэфирной связи сукцинил-КоА сопряжено с фосфорилированием гуанозиндифосфата (ГДФ):
Ф +ГДФ Сукцинат + ГТФ +КоА
Фосфорильная группа ГТФ легко переносится на АДФ с образованием АТФ:
ГТФ + АДФ АТФ + ГДФ
Это единственная реакция цикла, являющаяся реакцией субстратного фосфорилирования.
VIII. Это третья окислительно-восстановительная реакция:
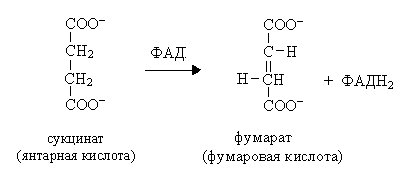
X 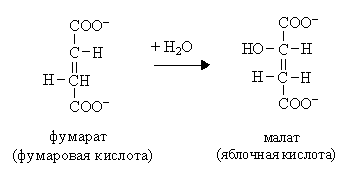 . Четвертая окислительно-восстановительная реакция:
. Четвертая окислительно-восстановительная реакция:
В цикле Кребса образуются углекислый газ, протоны, электроны. Четыре реакции цикла являются окислительно-восстановительными, катализируются ферментами – дегидрогеназами, содержащими коферменты НАД, ФАД. Коферменты захватывают образующиеся Н + и ē и передают их в дыхательную цепь (цепь биологического окисления). Элементы дыхательной цепи находятся на внутренней мембране митохондрий.
Тут вы можете оставить комментарий к выбранному абзацу или сообщить об ошибке.
Источник
4.1.2 Цикл Кребса. Механизмы регуляции цикла. Энергетическая эффективность процесса, значение
В анаэробных условиях пировиноградная кислота (пируват) подвергается дальнейшим превращениям в ходе спиртового, молочнокислого и других видов брожений, при этом NADH используется для восстановления конечных продуктов брожения, регенерируя в окисленную форму. Последнее обстоятельство поддерживает процесс гликолиза, для которого необходим окисленный NAD + . В присутствии достаточного количества кислорода пируват полностью окисляется до С02 и Н20 в дыхательном цикле, получившем название цикла Кребса или цикла ди- и трикарбоновых кислот. Все участки этого процесса локализованы в матриксе или во внутренней мембране митохондрий.
Последовательность реакций в цикле Кребса. Участие органических кислот в дыхании давно привлекало внимание исследователей. Еще в 1910 г. шведский химик Т. Тунберг показал, что в животных тканях содержатся ферменты, способные отнимать водород от некоторых органических кислот (янтарной, яблочной, лимонной). В 1935 г. А. Сент-Дьердьи в Венгрии установил, что добавление к измельченной мышечной ткани небольших количеств янтарной, фумаровой, яблочной или щавелевоуксуснсй кислот резко активирует поглощение тканью кислорода.
Учитывая данные Тунберга и Сент-Дьердьи и исходя из собственных экспериментов по изучению взаимопревращения различных органических кислот и их влияния на дыхание летательной мышцы голубя, английский биохимик Г. А. Кребс в 1937 г. предложил схему последовательности окисления ди- и трикарбоновых кислот до С02 через «цикл лимонной кислоты» да счет отнятия водорода. Этот цикл и был назван его именем.
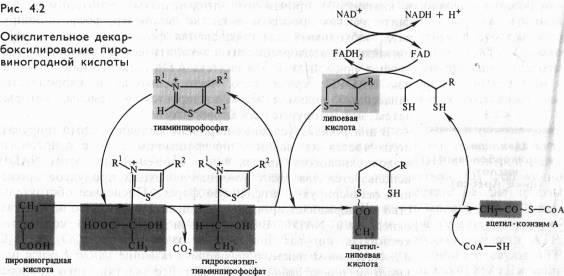
Непосредственно в цикле окисляется ле сам пируват, а его производное — ацетил-СоА. Таким образом, первым этапом на пути окислительного расщепления ПВК является процесс образования активного ацетила в ходе окислительного декарбоксилирования. Окислительное декарбоксилирование пирувата осуществляется при участии пируватдегидрогеназного мультифёрментного комплекса. В состав его входят три фермента и пять коферментов. Коферментами служат тиаминпирофосфат (ТПФ) — фосфорилированное производное витамина Вь липоевая кислота, коэнзим A, FAD и NAD + . Пируват взаимодействует с ТПФ (декарбоксилазой), при этом отщепляется С02 и образуется гидроксиэтильное производное ТПФ (рис. 4.2). Последнее вступает в реакцию с окисленной формой липоевой кислоты. Дисульфидная связь липоевой кислоты разрывается и происходит окислительно-восстановительная реакция: гидроксиэтильная группа, присоединенная к одному атому серы, окисляется в ацетильную (при этом возникает высокоэнергетическая тиоэфирная связь), а другой атом серы липоевой кислоты восстанавливается. Образовавшаяся ацетиллипоевая кислота взаимодействует с коэнзимом А, возникают ацетил- СоА и восстановленная форма липоевой кислоты. Водород липоевой кислоты переносится затем на FAD и далее на NAD + . В результате окислительного декарбоксилирования пирувата образуются ацетил-СоА, С02 и NADH.
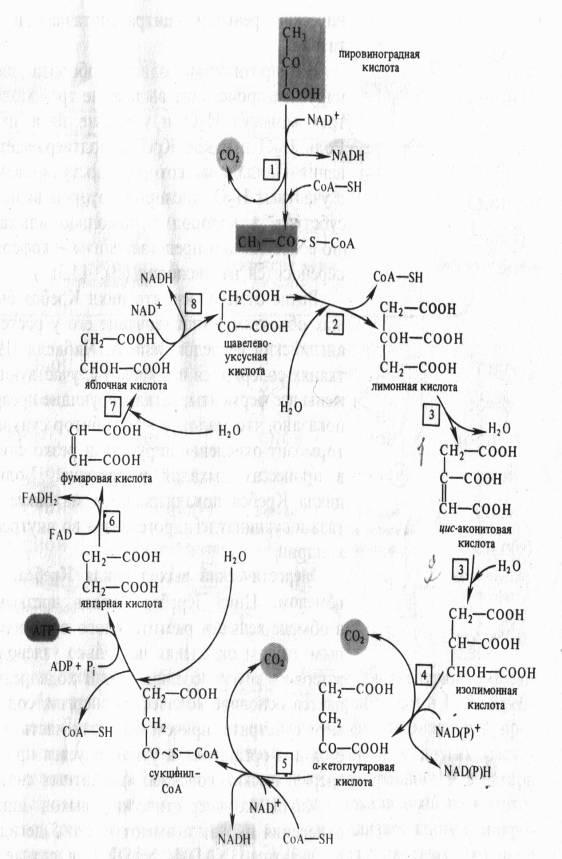
Дальнейшее окисление ацетил-СоА осуществляется в ходе циклического процесса. Цикл Кребса начинается с взаимодействия ацетил-СоА с енольной формой щавелевоуксусной кислоты. В этой реакции под действием фермента цитратсинтазы образуется лимонная кислота. Следующий этап цикла включает две реакции и катализируется ферментом аконитазой, или аконитатгидратазой. В первой реакции в результате дегидратации лимонной кислоты образуется цис-аконитовая. Во второй реакции аконитат гидратируется и синтезируется изолимонная кислота. Изолимонная кислота под действием NAD- или NADP-зависимой изоцитратдегидрогеназы окисляется в нестойкое соединение — щавелевоянтарную кислоту, которая тут же декарбоксилируется с образованием а-кетоглутаровой кислоты (а-оксоглутаровой кислоты).
а-Кетоглутарат, подобно пирувату, подвергается реакции окислительного декарбоксилирования. а-Кетоглутаратдегидрогеназный мультиэнзимный комплекс сходен с рассмотренным выше пируватдегидрогеназным комплексом. В ходе реакции окислительного декарбоксилирования а-кетоглутарата выделяется С02, образуются NADH и сукцинил-СоА.
Подобно ацетил-СоА, сукцинил-СоА является высокоэнергетическим тиоэфиром. Однако если в случае с ацетил-СоА энергия тиоэфирной связи расходуется на синтез лимонной кислоты, энергия сукцинил-CoA может трансформироватся в образование фосфатной связи АТР. При участии сукцинил- СоА-синтетазы из сукцинил-СоА, ADP и Н3Р04 образуются янтарная кислота (сукцинат), АТР, регенерирует молекула СоА. АТР образуется в результате субстратного фосфорилирования.
На следующем этапе янтарная кислота окисляется до фумаровой. Реакция катализируется сукцинатдегидрогеназой, коферментом которой является FAD. Фумаровая кислота под действием фумаразы или фумаратгидратазы, присоединяя Н20, превращается в яблочную кислоту (малат). И, наконец, на последнем этапе цикла яблочная кислота с помощью NAD- зависимой малатдегидрогеназы окисляется в щавелевоуксусную. ЩУК, которая самопроизвольно переходит в енольную форму, реагирует с очередной молекулой ацетил-СоА и цикл повторяется снова.
Следует отметить, что большинство реакций цикла обратимы, однако ход цикла в целом практически необратим. Причина этого в том, что в цикле есть две сильно экзергонические реакции — цитратсинтазная и сукцинил-СоА-синтетазная.
На протяжении одного оборота цикла при окислении пирувата происходит выделение трех молекул С02, включение трех молекул Н2О и удаление пяти пар атомов водорода. Роль Н2О в цикле Кребса подтверждает правильность уравнения Палладина, который постулировал, что дыхание идет с участием Н2О, кислород которой включается в окисляемый субстрат, а водород с помощью «дыхательных пигментов» (по современным представлениям — коферментов дегидрогеназ) переносится на кислород .
Выше отмечалось, что цикл Кребса был открыт на животных объектах. Существование его у растений впервые доказал английский исследователь А. Чибнелл (1939).В растительных тканях содержатся все кислоты, участвующие в цикле; обнаружены все ферменты, катализирующие превращение этих кислот; показано, что малонат — ингибитор сункцинатдегидрогеназы — тормозит окисление пирувата и резко снижает поглощение 02 в процессах дыхания у растений. Большинство ферментов цикла Кребса
локализовано в матриксе митохондрий, аконитаза и сукцинатдегидрогеназа — во внутренней мембране митохондрии.
Энергетический выход цикла Кребса, его связь с азотным обменом. Цикл Кребса. играет чрезвычайно важную роль в обмене веществ растительного организма. Он служит конечным этапом окисления не только углеводов, но также белков, жиров и других соединений. В ходе реакций цикла освобождается основное количество энергии, содержащейся в окисляемом субстрате, причем большая часть этой энергии не теряется для организма, а утилизируется при образовании высокоэнергетических конечных фосфатных связей АТР.
Каков же энергетический выход цикла Кребса? В ходе окисления пирувата имеют место 5 дегидрирований, при этом получаются 3NADH, NADPH (в случае изоцитратдегидрогеназы) и FADH2. Окисление каждой молекулы NADH (NADPH) при участии компонентов электронтранспортной цепи митохондрий дает по 3 молекулы АТР, а окисление FADH2 — 2АТР. Таким образом при полном окислении пирувата образуются 14 молекул АТР. Кроме того, 1 молекула АТР синтезируется ; в цикле Кребса в ходе субстратного фосфорилирования. Следовательно, при окислении одной молекуйы пирувата может образоваться 15 молекул АТР. А поскольку в процессе гликолиза из молекулы глюкозы возникают две молекулы пирувата, их окисление даст 30 молекул АТР.
Итак, при окислении глюкозы в процессе дыхания при функционировании гликолиза и цикла Кребса в общей сложности образуются 38 молекул АТР (8 АТР связаны с глико- лизом). Если принять, что энергия третьей сложноэфирнои фосфатной связи АТР равняется 41,87 кДж/моль (10 ккал/моль), то энергетический выход гликолитического пути аэробного дыхания составляет 1591 кДж/моль (380 ккал/моль).
Регуляция цикла Кребса. Дальнейшее использование образующегося из пирувата ацетил-СоА зависит от энергетического состояния клетки. При малой энергетической потребности клетки дыхательным контролем тормозится работа дыхательной цепи, а следовательно, реакций ЦТК и образования интермедиатов цикла, в том числе оксалоацетата, вовлекающего ацетил-СоА в цикл Кребса. Это приводит к большему использованию ацетил-СоА в синтетических процессах, которые также потребляют энергию.
Особенностью регуляции ЦТК является зависимость всех четырех дегидрогеназ цикла (изоцитратдегидрогеназы, а-кетоглутаратдегидрогеназы, сукцинатдегидрогеназы, малатдегидрогеназы) от отношения [NADH]/[NAD + ]. Активность цитратсинтазы тормозится высокой концентрацией АТР и собственным продуктом — цитратом. Изоцитратдегидрогеназа ингибируется NADH и активируется цитратом. а-Кето- глутаратдегидрогеназа подавляется продуктом реакции — сукцинил-СоА и активируется аденилатами. Окисление сукцината сукцинатдегидрогеназой тормозится оксалоацетатом и ускоряется АТР, ADP и восстановленным убихиноном (QH2). Наконец, малатдегидрогеназа ингибируется оксалоацетатом и у ряда объектов — высоким уровнем АТР. Однако степень участия величины энергетического заряда, или уровня адениновых нуклеотидов, в регуляции активности цикла Кребса у растений до конца не выяснена.
Регулирующую роль может играть также альтернативный путь транспорта электронов в растительных митохондриях. В условиях высокого содержания АТР, когда активность основной дыхательной цепи снижена, окисление субстратов через альтернативную оксидазу (без образования АТР) продолжается, что поддерживает на низком уровне отношение NADH/NAD + и снижает уровень АТР. Все это позволяет циклу Кребса функционировать.
Источник