ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ
Установлено, что окисление жирных кислот протекает в печени, почках, скелетных и сердечной мышцах, в жировой ткани. В мозговой ткани скорость окисления жирных кислот весьма незначительна; основным источником энергии в мозговой ткани служит глюкоза.
В 1904 г. Ф. Кнооп (F. Knoop) выдвинул гипотезу β-окисления жирных кислот на основании опытов по скармливанию собакам различных жирных кислот, в которых один атом водорода в концевой метильной группе (ω-углеродного атома) был замещен радикалом (С6Н5–).
Ф. Кнооп высказал предположение, что окисление молекулы жирной кислоты в тканях организма происходит в β-положении. В результате от молекулы жирной кислоты последовательно отщепляются двууглеродные фрагменты со стороны карбоксильной группы.
Жирные кислоты, входящие в состав естественных жиров животных и растений, имеют четное число углеродных атомов. Любая такая кислота, от которой отщепляется по паре углеродных атомов, в конце концов проходит через стадию масляной кислоты. После очередного β-окисления масляная кислота становится ацетоуксусной. Последняя затем гидроли-зуется до двух молекул уксусной кислоты. Теория β-окисления жирных кислот, предложенная Ф. Кноопом, в значительной мере послужила основой современных представлений о механизме окисления жирных кислот.
Доставка жирных кислот к месту их окисления – к митохондриям – происходит сложным путем: при участии альбумина осуществляется транспорт жирной кислоты в клетку; при участии специальных белков (fatty acid binding proteins, FABP) – транспорт в пределах цитозоля; при участии карнитина – транспорт жирной кислоты из цитозоля в митохондрии.
Процесс окисления жирных кислот складывается из следующих основных этапов.
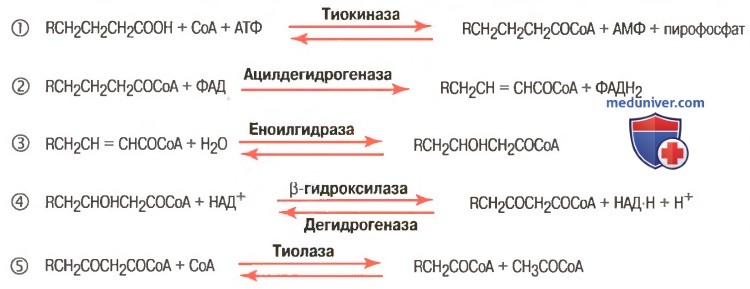
Активация жирных кислот. Свободная жирная кислота независимо от длины углеводородной цепи является метаболически инертной и не может подвергаться никаким биохимическим превращениям, в том числе окислению, пока не будет активирована. Активация жирной кислоты протекает на наружной поверхности мембраны митохондрий при участии АТФ, коэнзима A (HS-KoA) и ионов Mg 2+ . Реакция катализируется ферментом ацил-КоА-синтетазой:
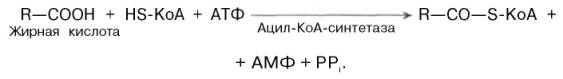
В результате реакции образуется ацил-КоА, являющийся активной формой жирной кислоты.
Считают, что активация жирной кислоты протекает в 2 этапа. Сначала жирная кислота реагирует с АТФ с образованием ациладенилата, представляющим собой эфир жирной кислоты и АМФ. Далее сульфгидрильная группа КоА действует на прочно связанный с ферментом ациладенилат с образованием ацил-КоА и АМФ.
Транспорт жирных кислот внутрь митохондрий. Коэнзимная форма жирной кислоты, в равной мере как и свободные жирные кислоты, не обладает способностью проникать внутрь митохондрий, где, собственно, и протекает их окисление. Переносчиком активированных жирных кислот
с длинной цепью через внутреннюю митохондриальную мембрану служит карнитин. Ацильная группа переносится с атома серы КоА на гидроксильную группу карнитина с образованием ацилкарнитина, который диффундирует через внутреннюю митохондриальную мембрану:
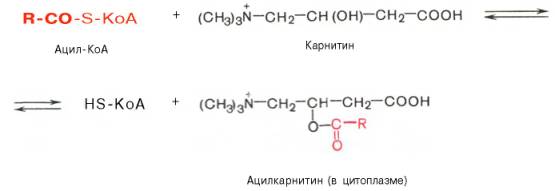
Реакция протекает при участии специфического цитоплазматического фермента карнитин-ацилтрансферазы. Уже на той стороне мембраны, которая обращена к матриксу, ацильная группа переносится обратно на КоА, что термодинамически выгодно, поскольку О-ацильная связь в кар-нитине обладает высоким потенциалом переноса группы. Иными словами, после прохождения ацилкарнитина через мембрану митохондрий происходит обратная реакция – расщепление ацилкарнитина при участии HS-KoA и митохондриальной карнитин-ацилтрансферазы:
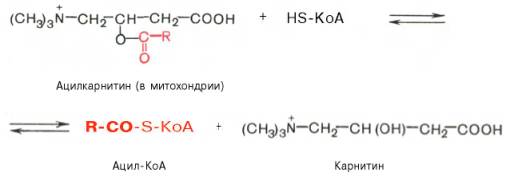
Внутримитохондриальное окисление жирных кислот. Процесс окисления жирной кислоты в митохондриях клетки включает несколько последовательных энзиматических реакций.
Первая стадия дегидрирования. Ацил-КоА в митохондриях прежде всего подвергается ферментативному дегидрированию, при этом ацил-КоА теряет 2 атома водорода в α- и β-положениях, превращаясь в КоА-эфир ненасыщенной кислоты. Таким образом, первой реакцией в каждом цикле распада ацил-КоА является его окисление ацил-КоА-де-гидрогеназой, приводящее к образованию еноил-КоА с двойной связью между С-2 и С-3:
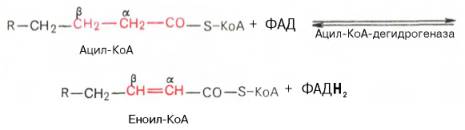
Существует несколько ФАД-содержащих ацил-КоА-дегидрогеназ, каждая из которых обладает специфичностью по отношению к ацил-КоА с определенной длиной углеродной цепи.
Стадия гидратации. Ненасыщенный ацил-КоА (еноил-КоА) при участии фермента еноил-КоА-гидратазы присоединяет молекулу воды. В результате образуется β-оксиацил-КоА (или 3-гидроксиацил-КоА):
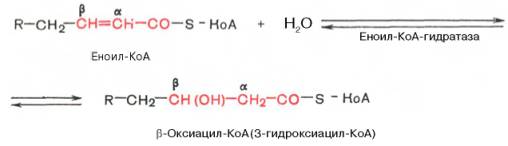
Заметим, что гидратация еноил-КоА стереоспецифична, подобно гидратации фумарата и аконитата (см. с. 348). В результате гидратации транс-Δ 2 -двойной связи образуется только L-изомер 3-гидроксиацил-КоА.
Вторая стадия дегидрирования. Образовавшийся β-оксиацил-КоА (3-гидроксиацил-КоА) затем дегидрируется. Эту реакцию катализируют НАД + -зависимые дегидрогеназы:
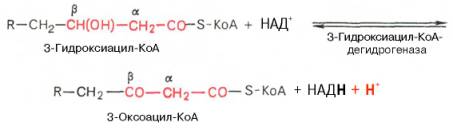
Тиолазная реакция. В ходе предыдущих реакций происходило окисление метиленовой группы при С-3 в оксогруппу. Тиолазная реакция представляет собой расщепление 3-оксоацил-КоА с помощью тиоловой группы второй молекулы КоА. В результате образуется укороченный на два углеродных атома ацил-КоА и двууглеродный фрагмент в виде ацетил-КоА. Данная реакция катализируется ацетил-КоА-ацилтрансферазой (β-ке-тотиолазой):
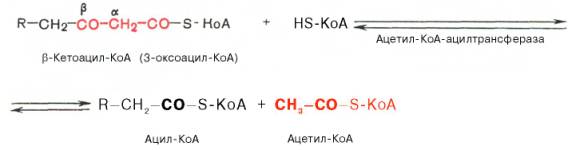
Образовавшийся ацетил-КоА подвергается окислению в цикле трикар-боновых кислот, а ацил-КоА, укоротившийся на два углеродных атома, снова многократно проходит весь путь β-окисления вплоть до образования бутирил-КоА (4-углеродное соединение), который в свою очередь окисляется до 2 молекул ацетил-КоА (рис. 11.2). Например, при окислении пальмитиновой кислоты (С16) повторяется 7 циклов β-окисления. Запомним, что при окислении жирной кислоты, содержащей п углеродных
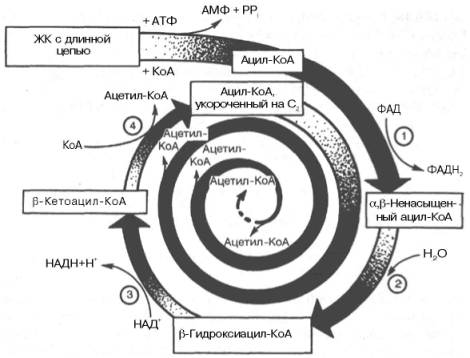
Рис. 11.2. Окисление жирной кислоты («спираль Линена»). Подробно представлен первый цикл окисления — укорочение цепи жирной кислоты на два углеродных атома. Остальные циклы аналогичны первому (по А.Н. Климову и Н.Г. Никульче-вой).
1 — ацил-КоА-дегидрогеназа (КФ 1.3.99.3) ; 2 — еноил-КоА-гидратаза (КФ 4.2.1.17.); 3 — β-гидро-ксиацил-КоА-дегидрогеназа (КФ 1.1.1.35); 4 — тиолаза (КФ 2.3.1.9).
атомов, происходит n/2–1 цикл β-окисления (т.е. на один цикл меньше, чем n/2, так как при окислении бутирил-КоА сразу происходит образование 2 молекул ацетил-КоА) и всего получится п/2 молекул ацетил-КоА. Следовательно, суммарное уравнение β-окисления активированной кислоты можно записать так:
Пальмитоил-КоА + 7ФАД + 7НАД + + 7Н2O + 7HS-KoA –>
–> 8Ацетил-КоА + 7ФАДН2 + 7НАДН + 7Н + .
Баланс энергии. При каждом цикле β-окисления образуются одна молекула ФАДН2 и одна молекула НАДН. Последние в процессе окисления в дыхательной цепи и сопряженного с ним фосфорилирования дают: ФАДН2 – 2 молекулы АТФ и НАДН – 3 молекулы АТФ, т.е. в сумме за один цикл образуется 5 молекул АТФ. При окислении пальмитиновой кислоты образуется 5 х 7 = 35 молекул АТФ. В процессе β-окисления пальмитиновой кислоты образуется 8 молекул ацетил-КоА, каждая из которых, «сгорая» в цикле трикарбоновых кислот, дает 12 молекул АТФ, а 8 молекул ацетил-КоА дадут 12 х 8 = 96 молекул АТФ.
Таким образом, всего при полном β-окислении пальмитиновой кислоты образуется 35 + 96 = 131 молекула АТФ. С учетом одной молекулы АТФ, потраченной в самом начале на образование активной формы пальмитиновой кислоты (пальмитоил-КоА), общий энергетический выход при полном окислении одной молекулы пальмитиновой кислоты в условиях животного организма составит 131 – 1 = 130 молекул АТФ. Изменение свободной энергии ΔF при полном сгорании 1 моля пальмитиновой кислоты составляет 2338 ккал, а богатая энергией фосфатная связь АТФ характеризуется величиной 7,6 ккал/моль. Нетрудно подсчитать, что примерно 990 ккал (7,6 х 130), или 42% от всей потенциальной энергии пальмитиновой кислоты при ее окислении в организме, используется для ресинтеза АТФ, а оставшаяся часть, очевидно, теряется в виде тепла.
Следовательно, эффективность накопления энергии в результате окисления жирных кислот при стандартных условиях составляет
Источник
Бета окислении высших жирных кислот участвует витамин
а) Расщепление жирных кислот до ацетилкоэнзима А путем бета-окисления. Молекула жирной кислоты расщепляется в митохондрии путем постепенного отщепления двууглеродных фрагментов в виде ацетилкоэнзима А (ацетил-КоА). Этот процесс, ход которого представлен на рисунке ниже, называют процессом бета-окисления.
 Бета-окисление жирных кислот в результате дает ацетилкоэнзим А
Бета-окисление жирных кислот в результате дает ацетилкоэнзим А
Обратите внимание, что первый этап бета-окисления представляет собой взаимодействие молекулы жирной кислоты с коэнзимом А (КоА) с образованием ацил-КоА жирной кислоты. В уравнениях 2, 3 и 4 бета-углерод (второй углерод справа) ацил-КоА жирной кислоты взаимодействует с молекулой кислорода, вследствие этого бета-углерод окисляется.
В правой части уравнения 5 двууглеродная часть молекулы отщепляется, образуя ацетил-КоА, выделяющийся во внеклеточную жидкость. В то же время другая молекула КоА взаимодействует с концом оставшейся части молекулы жирной кислоты, вновь формируя ацил-КоА жирной кислоты. Сама молекула жирной кислоты в это время становится короче на 2 атома углерода, т.к. первый ацетил-КоА уже отделился от ее терминали.
Затем эта укоротившаяся молекула ацил-КоА жирной кислоты выделяет еще 1 молекулу ацетил-КоА, что приводит к укорочению исходной молекулы жирной кислоты еще на 2 атома углерода. Кроме высвобождения молекул ацетил-КоА из молекул жирных кислот в ходе этого процесса выделяются 4 атома углерода.
б) Окисление ацетил-КоА. Образующиеся в митохондриях в ходе процесса бета-окисления жирных кислот молекулы ацетил-КоА немедленно поступают в цикл лимонной кислоты и, взаимодействуя прежде всего с щавелево-уксусной кислотой, образуют лимонную кислоту, которая затем последовательно окисляется посредством хемоосмотическои. системы окисления митохондрий. Чистый выход реакции цикла лимонной кислоты на 1 молекулу аце-тил-КоА составляет:

Таким образом, после начального расщепления жирной кислоты с образованием ацетил-КоА окончательное их расщепление осуществляется так же, как расщепление ацетил-КоА, образовавшегося из пировиноградной кислоты в процессе метаболизма глюкозы. Образующиеся при этом атомы водорода окисляются той же системой окисления митохондрий, которая используется в процессе окисления углеводов, с образованием большого количества аденозинтрифосфата.
в) При окислении жирных кислот образуется огромное количество АТФ. На рисунке выше показано, что 4 атома водорода, высвобождающиеся при отделении ацетил-КоА от цепочки жирной кислоты, выделяются в виде ФАДН2, НАД-Н и Н+, поэтому при расщеплении 1 молекулы стеариновой кислоты образуется, кроме 9 молекул ацетил-КоА, еще 32 атома водорода. В процессе расщепления каждой из 9 молекул ацетил-КоА в цикле лимонной кислоты выделяются еще 8 атомов водорода, что в итоге дает 72 атома водорода.
Суммарно при расщеплении 1 молекулы стеариновой кислоты выделяются 104 атома водорода. Из этого общего количества 34 атома выделяются, будучи связанными с флавопротеинами, а остальные 70 высвобождаются в форме, связанной с никотинамидадениндинуклеотидом, т.е. в виде НАД-Н+ и Н+.
Окисление водорода, связанного с этими двумя типами веществ, осуществляется в митохондриях, но они вступают в процесс окисления в разных точках, поэтому окисление каждого из 34 атомов водорода, связанных с флавопротеинами, приводит к выделению 1 молекулы АТФ. Еще 1,5 молекулы АТФ синтезируется из каждых 70 НАД+ и Н+. Это дает к 34 еще 105 молекул АТФ (т.е. всего 139) при окислении водорода, отщепляющегося при окислении каждой молекулы стеариновой кислоты.
Дополнительно 9 молекул АТФ образуются в цикле лимонной кислоты (помимо АТФ, получаемой при окислении водорода), по 1 на каждую из 9 молекул метаболизирующегося ацетил-КоА. Итак, при полном окислении 1 молекулы стеариновой кислоты образуются в сумме 148 молекул АТФ. С учетом того, что при взаимодействии стеариновой кислоты с КоА на начальной стадии метаболизма этой жирной кислоты расходуются 2 молекулы АТФ, чистый выход АТФ составляет 146 молекул.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
— Вернуться в оглавление раздела «Физиология человека.»
Источник
Бета-окисление – определение, шаги и тест
Определение бета-окисления
Бета-окисление – это метаболический процесс, включающий несколько стадий, посредством которых молекулы жирных кислот расщепляются для производства энергии. Более конкретно, бета-окисление состоит в том, чтобы долго разрушаться жирные кислоты которые были превращены в ацил-СоА-цепи в постепенно уменьшающиеся жирные ацил-КоА-цепи. Эта реакция высвобождает ацетил-КоА, FADH2 и NADH, три из которых затем вступают в другой метаболический процесс, называемый циклом лимонной кислоты или Цикл Кребса, в котором производится АТФ для использования в качестве энергии. Бета-окисление продолжается до тех пор, пока не образуются две молекулы ацетил-КоА, и цепь ацил-КоА полностью не разрушается. В эукариотических клетках бета-окисление происходит в митохондрии в то время как в прокариотических клетках это происходит в цитозоль.
Для того, чтобы произошло бета-окисление, жирные кислоты должны сначала попасть в клетка сквозь клеточная мембрана, а затем привязать к коэнзим A (CoA), образуя жирный ацил CoA и, в случае эукариотических клеток, попадает в митохондрии, где происходит бета-окисление.
Где происходит бета-окисление?
Бета-окисление происходит в митохондриях эукариотических клеток и в цитозоле прокариотических клеток. Однако, прежде чем это произойдет, жирные кислоты должны сначала проникнуть в клетку, а в случае эукариотических клеток – в митохондрии. В случаях, когда цепочки жирных кислот слишком длинные, чтобы проникнуть в митохондрии, бета-окисление также может происходить в пероксисомах.
Во-первых, белковые транспортеры жирных кислот позволяют жирным кислотам проходить через клеточную мембрану и проникать в цитозоль, поскольку отрицательно заряженные цепи жирных кислот не могут пересекать его иначе. Затем фермент жирная ацил-КоА-синтаза (или FACS) добавляет группу СоА к цепи жирных кислот, превращая ее в ацил-КоА.
В зависимости от длины цепь ацил-КоА будет проникать в митохондрии одним из двух способов:
Как упомянуто выше, если цепь ацил-КоА слишком длинная, чтобы обрабатываться в митохондриях, она будет разрушена в результате бета-окисления в пероксисомах. Исследования показывают, что очень длинные цепочки ацил-КоА разрушаются до тех пор, пока они не станут длиной 8 атомов углерода, после чего они транспортируются и вступают в цикл бета-окисления в митохондриях. Бета-окисление в пероксисомах дает H2O2 вместо FADH2 и NADH, в результате чего выделяется тепло.
Бета-этапы окисления
Бета-окисление происходит в четыре этапа: дегидрирование, гидратация, окисление и тиолизис. Каждый шаг катализируется отдельным ферментом.
Вкратце, каждый цикл этого процесса начинается с цепи ацил-КоА и заканчивается одной ацетил-КоА, одним FADH2, одним NADH и водой, и цепь ацил-КоА становится на два атома углерода короче. Общий выход энергии за цикл составляет 17 молекул АТФ (подробности о распаде см. Ниже). Этот цикл повторяется до тех пор, пока не образуются две молекулы ацетил-КоА, в отличие от одного ацил-КоА и одной ацетил-КоА. Четыре стадии бета-окисления описаны ниже, и их можно увидеть в ссылках на рисунки в конце каждого объяснения.
дегидрогенизация
На первом этапе ацил-КоА окисляется ферментом ацил-КоА-дегидрогеназой. Двойная связь образуется между вторым и третьим атомами углерода (С2 и С3) цепи ацил-КоА, входящей в цикл бета-окисления; конечным продуктом этой реакции является транс-2-еноил-КоА (транс-дельта-2-еноил-КоА). На этом этапе используется FAD и вырабатывается FADH2, который входит в цикл лимонной кислоты и образует АТФ для использования в качестве энергии. (Обратите внимание, что на следующем рисунке отсчет углерода начинается с правой стороны: самый правый углерод ниже атома кислорода – это С1, затем С2 слева, образующий двойную связь с С3, и т. Д.)
гидратация
На втором этапе двойная связь между С2 и С3 транс-Δ2-еноил-КоА гидратируется с образованием конечного продукта L-β-гидроксиацил-КоА, который имеет гидроксильная группа (ОН) в С2, вместо двойной связи. Эта реакция катализируется другим ферментом: еноил-КоА-гидратазой. Этот шаг требует воды.
оксидирование
На третьей стадии гидроксильная группа в С2 L-β-гидроксиацил-КоА окисляется NAD + в реакции, которая катализируется 3-гидроксиацил-КоА-дегидрогеназой. Конечными продуктами являются β-кетоацил-КоА и NADH + H. NADH входит в цикл лимонной кислоты и производит АТФ, который будет использоваться в качестве энергии.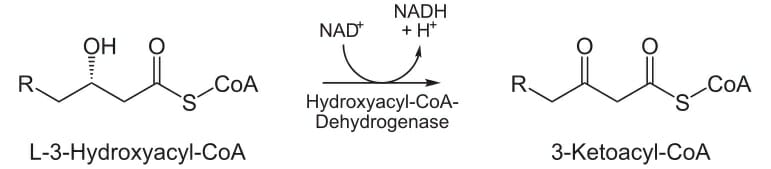
тиолиза
Наконец, на четвертом этапе β-кетоацил-КоА расщепляется тиольной группой (SH) другого КоА. молекула (КоА-SH). Фермент, который катализирует эту реакцию, является β-кетотиолазой. Расщепление происходит между С2 и С3; следовательно, конечные продукты представляют собой молекулу ацетил-КоА с исходными двумя первыми атомами углерода (С1 и С2) и цепью ацил-КоА на два атома углерода, которая короче, чем исходная цепь ацил-КоА, которая вступила в цикл бета-окисления.
Конец бета-окисления
В случае четных ацил-СоА-цепей бета-окисление заканчивается после того, как четырехуглеродная ацил-КоА-цепь распадается на две единицы ацетил-КоА, каждая из которых содержит два атома углерода. Молекулы ацетил-КоА вступают в цикл лимонной кислоты с образованием АТФ.
В случае нечетных цепей ацил-СоА бета-окисление происходит таким же образом, за исключением последнего этапа: вместо разрыва четырехцепочечной цепи ацил-КоА на две единицы ацетил-КоА, пятиуглеродный ацил -СоА цепь расщепляется на трехуглеродный пропионил-КоА и двухуглеродный ацетил-КоА. Другая химическая реакция затем превращает пропионил-КоА в сукцинил-КоА (см. Рисунок ниже), который входит в цикл лимонной кислоты с образованием АТФ.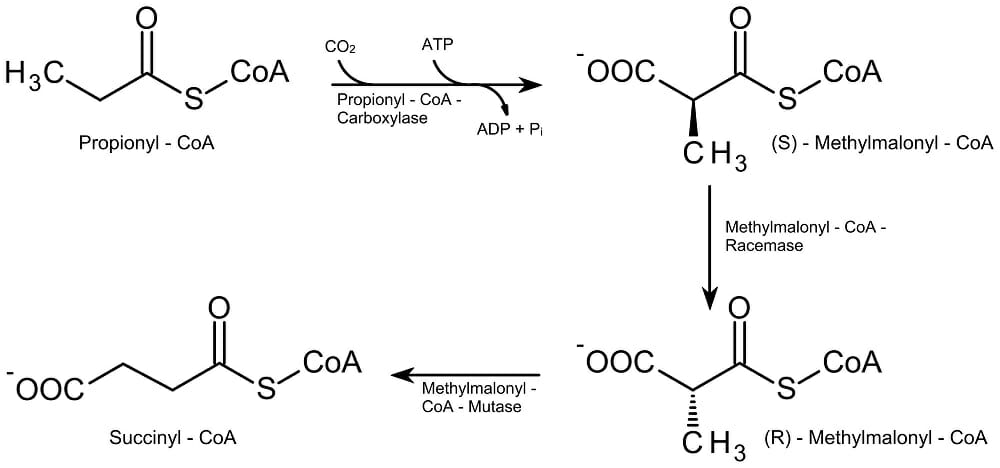
Выход энергии и конечные продукты
Каждый цикл бета-окисления дает 1 FADH2, 1 NADH и 1 ацетил-КоА, что по энергии эквивалентно 17 молекулам АТФ:
- 1 FADH2 (x 2 ATP) = 2 ATP
- 1 НАДН (х 3 АТФ) = 3 АТФ
- 1 ацетил-КоА (х 12 АТФ) = 12 АТФ
- Всего = 2 + 3 + 12 = 17 спс
Однако теоретический выход АТФ выше, чем реальный выход АТФ. В действительности в каждом цикле бета-окисления образуется эквивалент примерно 12-16 АТФ.
Помимо выхода энергии, жирная цепь ацил-CoaA становится короче на два атома углерода с каждым циклом. Кроме того, бета-окисление дает большое количество воды; это полезно для эукариотических организмов, таких как верблюды, учитывая их ограниченный доступ к питьевой воде.
викторина
1. Что делает бета-окисление?A. Расщеплять углеводы.B. Расщеплять белки.C. Расщепляют жирные кислоты.D. Расщепляют жирные кислоты и белки.D. Расщеплять углеводы и белки.
Ответ на вопрос № 1
С верно. Бета-окисление расщепляет жирные кислоты для производства энергии. В частности, жирные ацил-КоА-цепи расщепляются на ацетил-КоА, FADH2, NADH и воду.
2. Каков порядок этапов цикла бета-окисления?A. Дегидрирование, гидратация, окисление и тиолизис.B. Гидратация, дегидрирование, окисление и тиолизис.C. Дегидрирование, окисление, тиолиз и гидратация.D. Гидратация, дегидрирование, тиолиз и окисление
Ответ на вопрос № 2
верно. Стадии в порядке: дегидрирование, гидратация, окисление и тиолизис.
3. Какие конечные продукты дает каждый цикл бета-окисления?A. Одна ацил-КоА, одна NADH, вода и одна цепь ацетил-КоА на два атома углерода короче.B. Одна двухуглеродная ацил-КоА, одна NADH, вода и другая ацил-КоА цепь на два атома углерода короче.C. Одна ацил-КоА, одна FADH2, одна NADH, вода и одна цепь ацетил-КоА на два атома углерода короче.D. Одна ацетил-КоА, одна FADH2, одна NADH, вода и одна ацил-КоА цепь на два атома углерода короче.
Ответ на вопрос № 3
D верно. Эффективный цикл бета-окисления дает одну (двухуглеродную) ацетил-КоА, одну FADH2, одну NADH, воду и одну ацил-КоА-цепочку на два атома углерода. FADH2, NADH и ацетил-КоА позже вступают в цикл лимонной кислоты, что дает АТФ.
Источник