Ацетил-КоА карбоксилаза — Acetyl-CoA carboxylase
| Ацетил-КоА карбоксилаза | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Номер ЕС | 6.4.1.2 | ||||||||
| Количество CAS | 9023-93-2 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| BRENDA | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| Структуры PDB | RCSB PDB PDBe PDBsum | ||||||||
| Генная онтология | Amigo / QuickGO | ||||||||
| |||||||||
| Ацетил-КоА карбоксилаза альфа | |||||||
|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||
| Символ | ACACA | ||||||
| Альт. символы | ACAC, ACC1, ACCA | ||||||
| Ген NCBI | 31 год | ||||||
| HGNC | 84 | ||||||
| OMIM | 601557 | ||||||
| RefSeq | NM_198839 | ||||||
| UniProt | Q13085 | ||||||
| Прочие данные | |||||||
| Номер ЕС | 6.4.1.2 | ||||||
| Locus | Chr. 17 q21 | ||||||
| |||||||
| Ацетил-КоА карбоксилаза бета | |||||||
|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||
| Символ | ACACB | ||||||
| Альт. символы | ACC2, ACCB | ||||||
| Ген NCBI | 32 | ||||||
| HGNC | 85 | ||||||
| OMIM | 200350 | ||||||
| RefSeq | NM_001093 | ||||||
| UniProt | O00763 | ||||||
| Прочие данные | |||||||
| Номер ЕС | 6.4.1.2 | ||||||
| Locus | Chr. 12 q24.1 | ||||||
| |||||||
Ацетил-СоА — карбоксилазы ( АСС ) представляет собой биотина -зависимой фермент , который катализирует необратимую карбоксилирования из ацетил-КоА , чтобы произвести малонил-СоА за счет двух каталитических активностей, биотин — карбоксилазы (БК) и carboxyltransferase (КТ). АСС — это мультисубъединичный фермент у большинства прокариот и в хлоропластах большинства растений и водорослей, тогда как это большой мультидоменный фермент в цитоплазме большинства эукариот . Наиболее важной функцией ACC является обеспечение субстрата малонил-КоА для биосинтеза жирных кислот . Активностью ACC можно управлять на уровне транскрипции, а также с помощью модуляторов малых молекул и ковалентной модификации . Геном человека содержит гены двух разных ACC — ACACA и ACACB .
СОДЕРЖАНИЕ
Структура
Прокариоты и растения имеют мультисубъединичные АСС, состоящие из нескольких полипептидов. Активность биотинкарбоксилазы (BC), активность белка-носителя карбоксила биотина (BCCP) и активность карбоксилтрансферазы (CT), каждая, содержится в разных субъединицах. Стехиометрия этих субъединиц в холоферменте ACC различается у разных организмов. Люди и большинство эукариот развили ACC с каталитическими доменами CT и BC и доменами BCCP на единственном полипептиде. Большинство растений также имеют эту гомомерную форму в цитозоле. Функциональные области ACC, начинающиеся от N-конца до C-конца, представляют собой биотинкарбоксилазу (BC), связывающую биотин (BB), карбоксилтрансферазу (CT) и связывающую АТФ (AB). AB находится внутри BC. Биотин ковалентно присоединен через амидную связь к длинной боковой цепи лизина, находящегося в ВВ. Поскольку BB находится между участками BC и CT, биотин может легко перемещаться в оба активных центра, где это необходимо.
У млекопитающих, у которых экспрессируются две изоформы ACC, основным структурным различием между этими изоформами является удлиненный N-конец ACC2, содержащий направленную на митохондрии последовательность .

Субъединица биотинкарбоксилазы ацетил-КоА-карбоксилазы E. coli

Биотин-карбоксильная белковая субъединица ацетил-КоА-карбоксилазы E. coli
![]()
Субъединица карбоксилтрансферазы ацетил-КоА-карбоксилазы E. coli
Полипептиды, составляющие мультисубъединичные АСС прокариот и растений , кодируются разными генами. В кишечной палочки , АССА кодирует альфа — субъединицу карбоксилазы ацетил-СоА, и ACCD кодирует его бета — субъединицу.
Механизм
Общая реакция ACAC (A, B) протекает по двухступенчатому механизму. Первая реакция осуществляется до н.э. и включает в себя АТФ-зависимая карбоксилированию биотины с бикарбонатом , выступающим в качестве источника CO 2 . Карбоксильная группа переносится от биотина к ацетил-КоА с образованием малонил-КоА во второй реакции, которая катализируется ХТ.
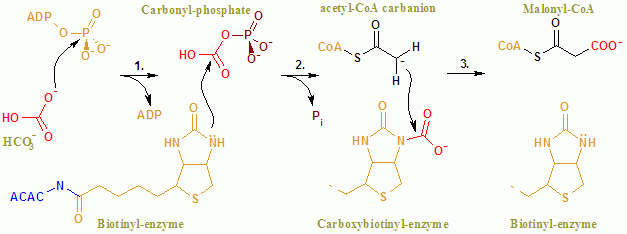
В активном центре реакция протекает при обширном взаимодействии остатков Glu296 и положительно заряженных Arg338 и Arg292 с субстратами. Два Mg 2+ координируются фосфатными группами АТФ и необходимы для связывания АТФ с ферментом. Бикарбонат депротонируется Glu296, хотя в растворе этот перенос протона маловероятен, поскольку pKa бикарбоната составляет 10,3. Фермент, по-видимому, манипулирует pKa, чтобы облегчить депротонирование бикарбоната. PKa бикарбоната снижается за счет его взаимодействия с положительно заряженными боковыми цепями Arg338 и Arg292. Кроме того, Glu296 взаимодействует с боковой цепью Glu211, взаимодействие, которое, как было показано, вызывает увеличение видимого pKa. После депротонирования бикарбоната кислород бикарбоната действует как нуклеофил и атакует гамма-фосфат на АТФ. Промежуточный карбоксифосфат быстро разлагается до CO 2 и PO 4 3– . PO 4 3- депротонирует биотин, создавая енолят, стабилизированный Arg338, который впоследствии атакует CO 2, что приводит к образованию карбоксибиотина. Карбоксибиотин перемещается в активный сайт карбоксилтрансферазы (СТ), где карбоксильная группа переносится на ацетил-КоА. В отличие от домена BC, о механизме реакции CT известно немного. Предлагаемый механизм — это высвобождение CO 2 из биотина, который впоследствии отрывает протон от метильной группы от ацетил-CoA-карбоксилазы. Полученный енолят атакует CO 2 с образованием малонил-КоА. В конкурирующем механизме отрыв протона согласован с атакой ацетил-КоА.
Функция
Функция ACC — регулировать метаболизм жирных кислот. Когда фермент активен, продукт, малонил-СоА, производится который представляет собой строительный блок для новых жирных кислот и может ингибировать передачу жирной ацильной группы из ацил — СоА , чтобы карнитин с карнитин ацилтрансферазы , который ингибирует бета-окисление в жирные кислоты в митохондриях .
У млекопитающих экспрессируются две основные изоформы ACC, ACC1 и ACC2, которые различаются как распределением в тканях, так и функцией. ACC1 находится в цитоплазме всех клеток, но он обогащен липогенной тканью, такой как жировая ткань и лактирующие молочные железы , где важен синтез жирных кислот. В окислительных тканях, таких как скелетные мышцы и сердце , соотношение экспрессируемого АСС2 выше. И ACC1, и ACC2 высоко экспрессируются в печени, где важны как окисление, так и синтез жирных кислот. Различия в распределении тканей указывают на то, что ACC1 поддерживает регуляцию синтеза жирных кислот, тогда как ACC2 в основном регулирует окисление жирных кислот (бета-окисление).
Регулирование

Регулирование ACC млекопитающих является сложным, чтобы контролировать два разных пула малонил-КоА, которые управляют либо ингибированием бета-окисления, либо активацией биосинтеза липидов.
ACC1 и ACC2 млекопитающих регулируются транскрипционно множеством промоторов, которые опосредуют изобилие ACC в ответ на состояние питания клеток. Активация экспрессии генов через разные промоторы приводит к альтернативному сплайсингу ; однако физиологическое значение конкретных изоферментов АЦК остается неясным. Чувствительность к статусу питания является результатом контроля этих промоторов факторами транскрипции, такими как белок 1 , связывающий регуляторный элемент стерола , который контролируется инсулином на уровне транскрипции, и ChREBP , экспрессия которого увеличивается при диете с высоким содержанием углеводов .
Через петлю прямой связи цитрат аллостерически активирует АСС. Цитрат может увеличить полимеризацию АСС для увеличения ферментативной активности; однако неясно, является ли полимеризация основным механизмом цитратного увеличения активности ACC или полимеризация является артефактом экспериментов in vitro. Другие аллостерические активаторы включают глутамат и другие дикарбоновые кислоты . Длинноцепочечные и короткоцепочечные жирные ацил-КоА являются ингибиторами АСС с отрицательной обратной связью.
Фосфорилирование может происходить, когда гормоны глюкагон или адреналин связываются с рецепторами клеточной поверхности , но основная причина фосфорилирования связана с повышением уровня АМФ при низком энергетическом статусе клетки, что приводит к активации АМФ-активируемой протеинкиназы. (АМПК). AMPK является основным регулятором киназы ACC, способным фосфорилировать ряд остатков серина на обеих изоформах ACC. На ACC1 AMPK фосфорилирует Ser79, Ser1200 и Ser1215. Протеинкиназа А также обладает способностью фосфорилировать АСС с гораздо большей способностью фосфорилировать АСС2, чем АСС1. Однако физиологическое значение протеинкиназы A в регуляции ACC в настоящее время неизвестно. Исследователи предполагают, что существуют другие киназы ACC, важные для его регуляции, поскольку существует множество других возможных сайтов фосфорилирования на ACC.
Когда инсулин связывается со своими рецепторами на клеточной мембране , он активирует фермент фосфатазу, называемый протеинфосфатазой 2A (PP2A), для дефосфорилирования фермента; тем самым снимая тормозящий эффект. Кроме того, инсулин индуцирует фосфодиэстеразу, которая снижает уровень цАМФ в клетке, таким образом ингибируя PKA, а также напрямую ингибирует AMPK.
Клинические последствия
На стыке путей синтеза и окисления липидов ACC представляет множество клинических возможностей для производства новых антибиотиков и разработки новых методов лечения диабета , ожирения и других проявлений метаболического синдрома . Исследователи стремятся использовать структурные различия между бактериальными и человеческими ACC для создания антибиотиков, специфичных для бактериальных ACC, чтобы минимизировать побочные эффекты для пациентов. Многообещающие результаты в отношении полезности ингибитора АСС включают открытие, что у мышей без экспрессии АСС2 наблюдается непрерывное окисление жирных кислот, сниженная масса тела и сниженная масса тела, несмотря на увеличение потребления пищи. Эти мыши также защищены от диабета. Недостаток ACC1 у мутантных мышей летален уже на эмбриональной стадии. Однако неизвестно, должны ли препараты, нацеленные на ACC у людей, быть специфичными для ACC2.
Фирсокостат (ранее GS-976, ND-630, NDI-010976) является мощным аллостерическим ингибитором ACC, действующим на BC-домен ACC. Фирсокостат разрабатывается фармацевтической компанией Gilead в 2019 году (фаза II) в рамках комбинированного лечения неалкогольного стеатогепатита (НАСГ), который считается все более частой причиной печеночной недостаточности.
Кроме того, селективные к растениям ингибиторы АЦК широко используются в качестве гербицидов , что предполагает клиническое применение против паразитов Apicomplexa, которые зависят от изоформы АЦК растительного происхождения, включая малярию .
Источник